
Whole Exome Sequencing Identifies PHF14 Mutations in Neurocytoma and Predicts Responsivity to the PDGFR Inhibitor Sunitinib
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
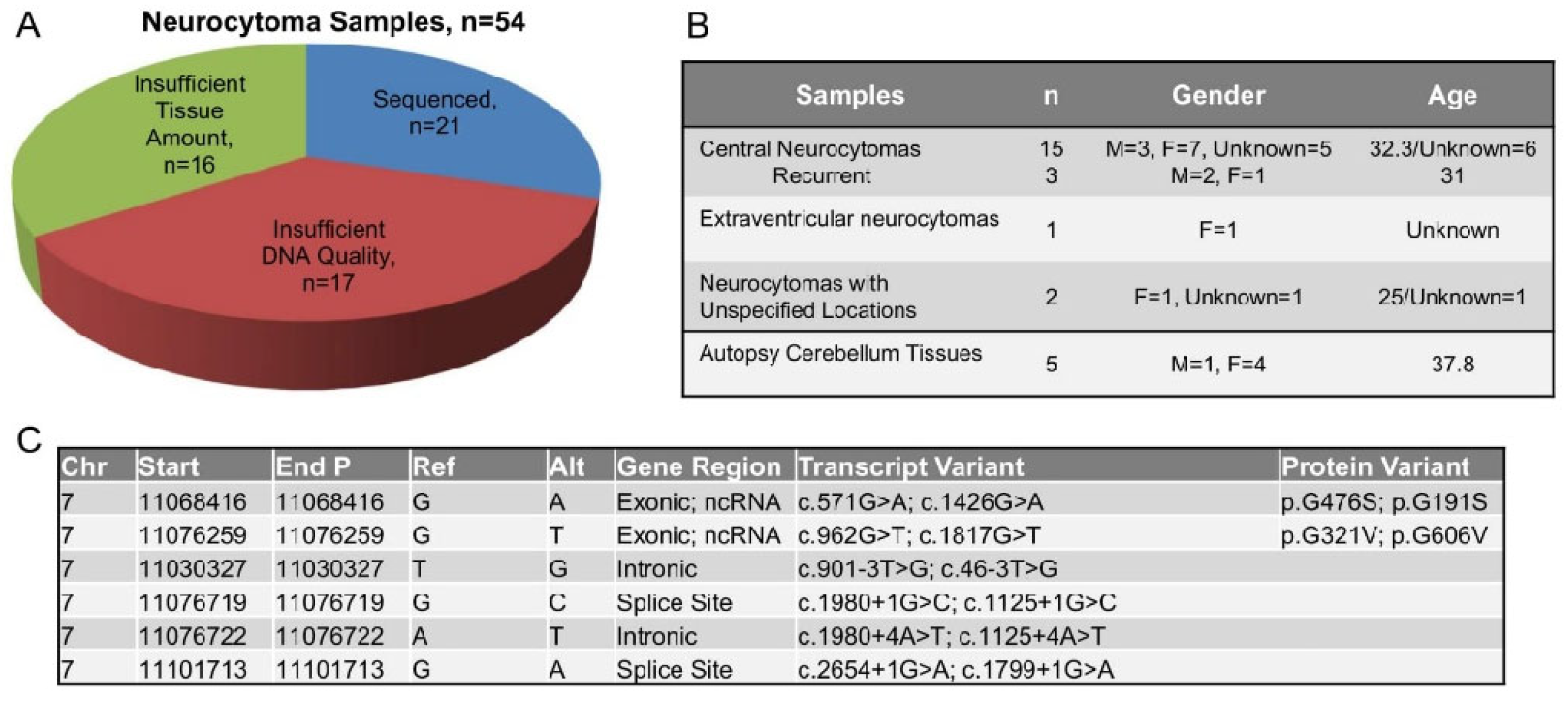
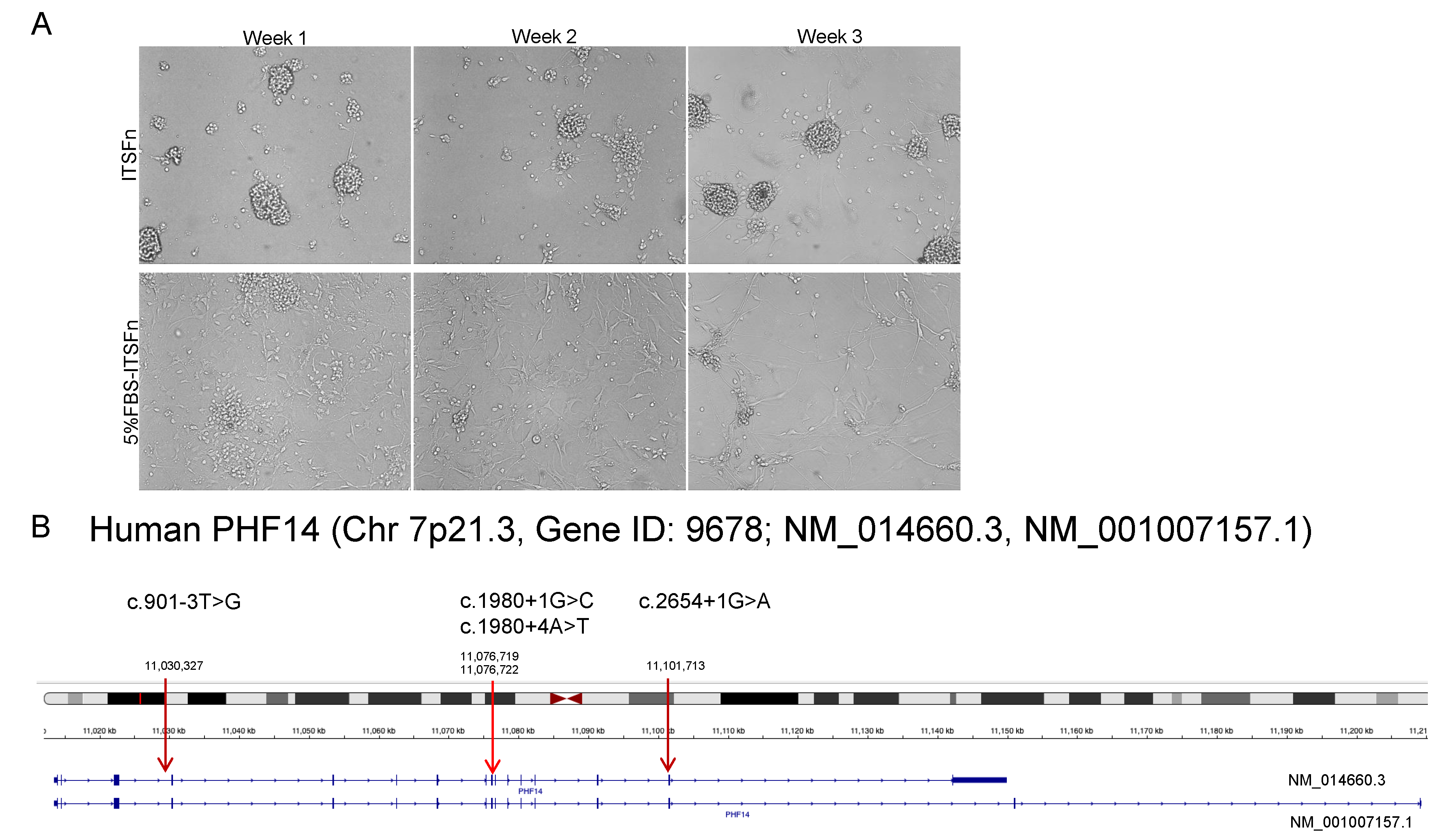
2.1. PHF14 Mutations in Human Neurocytoma
2.2. PHF14 Knockdown Enhances Cell Proliferation and Increases Anchorage Independent Cell Growth
2.3. PHF14 Depletion Increases PDGFRα Expression and Responsivity to Sunitinib
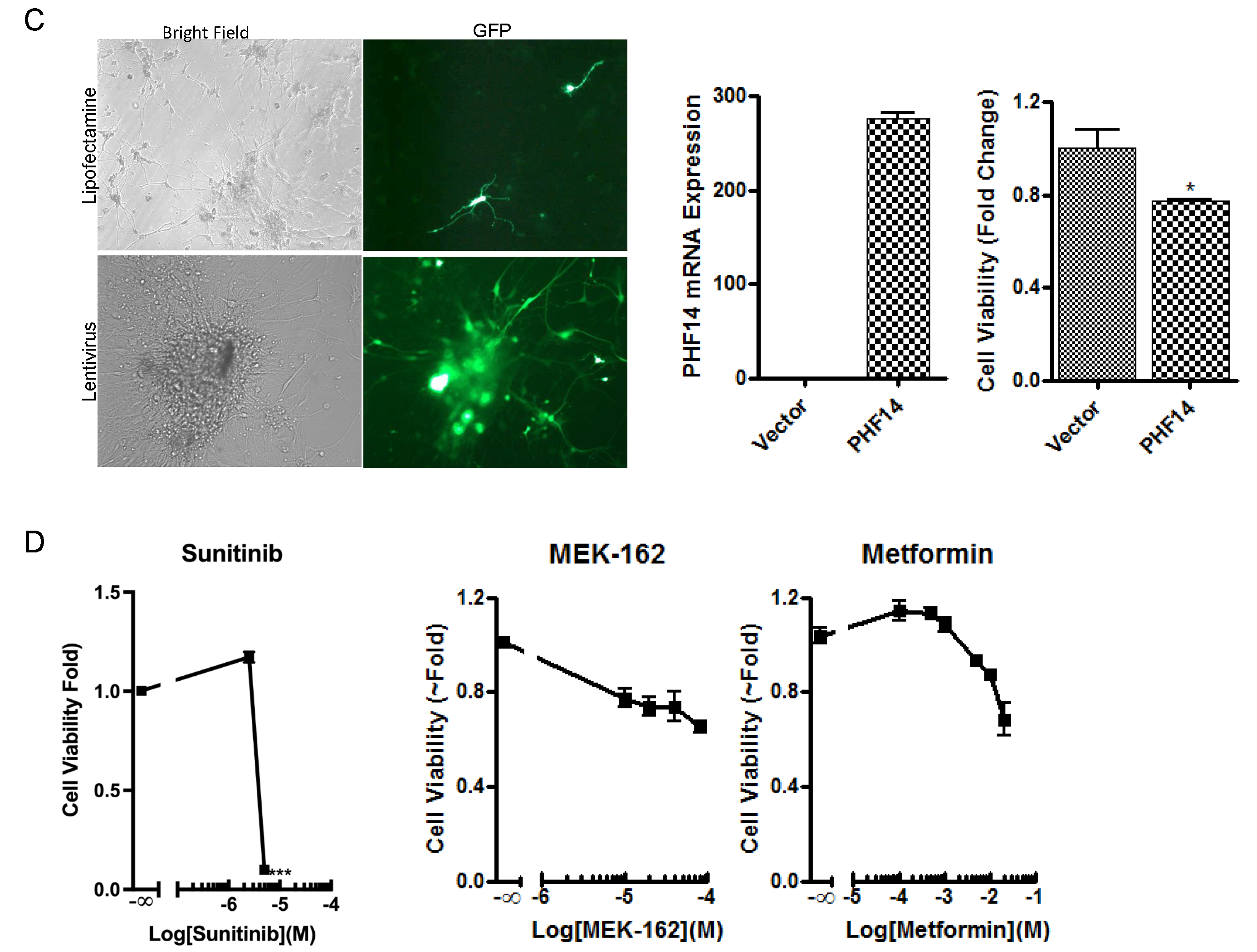
2.4. Overexpression of PHF14 Inhibits the Growth of Neurocytoma Primary Culture with PHF14 Defect
2.5. Confirmation of the Tumor Inhibitory Effect of PHF14 and Sunitinib In Vivo
3. Discussion
4. Materials and Methods
4.1. DNA Isolation and Whole Exome Sequencing
4.2. Bioinformatic Analysis of WES Data
4.3. Cell Culture and Reagents
4.4. RNA Microarray Processing and Bioinformatic Analysis
4.5. Human Neurocytoma Primary Cultures
4.6. Plasmid Constructs, Transfection and Viral Transduction
4.7. Cell Proliferation Assay
4.8. Anchorage-Independent Growth Assay
4.9. Real-Time PCR
4.10. Western Blotting
4.11. Tumor Xenograft Model
4.12. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Song, Y.; Kang, X.; Cao, G.; Li, Y.; Zhou, X.; Tong, Y.; Wang, W. Clinical characteristics and prognostic factors of brain central neurocytoma. Oncotarget 2016, 7, 76291–76297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, M.; Joo, J.-D.; Kim, B.-H.; Choe, G.; Kim, C.-Y. Hypothalamic Extraventricular Neurocytoma (EVN) in a Pediatric Patient: A Case of EVN Treated with Subtotal Removal Followed by Adjuvant Radiotherapy. Brain Tumor Res. Treat. 2016, 4, 35–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaji, H.; Saito, O.; Amano, S.; Kasahara, M.; Baba, S.; Namba, H. Extraventricular neurocytoma of the sellar region with spinal dissemination. Brain Tumor Pathol. 2012, 31, 51–56. [Google Scholar] [CrossRef]
- Kim, S.R.; Asa, S.L.; Heaney, A.P. Severe Hyponatremia in a Patient with Vasopressin-Producing Sellar Neurocytoma. In Proceedings of the Late-Breaking Tumor Biology III Session at the 97th Annual Meeting of the Endocrine Society, San Diego, CA, USA, 5–8 March 2015. LBS-059. [Google Scholar]
- Lee, S.J.; Bui, T.T.; Chen, C.H.J.; Lagman, C.; Chung, L.K.; Sidhu, S.; Seo, D.J.; Yong, W.H.; Siegal, T.L.; Kim, M.; et al. Central Neurocytoma: A Review of Clinical Management and Histopathologic Features. Brain Tumor Res. Treat. 2016, 4, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-L.; Shen, C.-C.; Wang, J.; Lu, C.-H.; Lee, H.-T. Central neurocytoma: A clinical, radiological and pathological study of nine cases. Clin. Neurol. Neurosurg. 2008, 110, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-D.; Li, W.-B.; Feng, J.; Qiu, X.-G. Long-term outcomes of adjuvant radiotherapy after surgical resection of central neurocytoma. Radiat. Oncol. 2014, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Korshunov, A.; Sycheva, R.; Golanov, A. Recurrent cytogenetic aberrations in central neurocytomas and their biological relevance. Acta Neuropathol. 2006, 113, 303–312. [Google Scholar] [CrossRef]
- Sander, C.; Wallenborn, M.; Brandt, V.P.; Ahnert, P.; Reuschel, V.; Eisenlöffel, C.; Krupp, W.; Meixensberger, J.; Holland, H. Central neurocytoma: SNP array analyses, subtel FISH, and review of the literature. Pathol.-Res. Pract. 2019, 215, 152397. [Google Scholar] [CrossRef]
- Sievers, P.; Stichel, D.; Schrimpf, D.; Sahm, F.; Koelsche, C.; Reuss, D.E.; Wefers, A.K.; Reinhardt, A.; Huang, K.; Ebrahimi, A.; et al. FGFR1:TACC1 fusion is a frequent event in molecularly defined extraventricular neurocytoma. Acta Neuropathol. 2018, 136, 293–302. [Google Scholar] [CrossRef]
- Aghajan, Y.; Malicki, D.M.; Levy, M.L.; Crawford, J.R. Atypical central neurocytoma with novel EWSR1-ATF1 fusion and MUTYH mutation detected by next-generation sequencing. BMJ Case Rep. 2019, 12, 226455. [Google Scholar] [CrossRef]
- Sanchez, R.; Zhou, M.-M. The PHD finger: A versatile epigenome reader. Trends Biochem. Sci. 2011, 36, 364–372. [Google Scholar] [CrossRef] [Green Version]
- Morrison, E.A.; Musselman, C.A. Chromatin Signaling and Diseases; Academic Press Inc.: Cambridge, MA, USA, 2016; pp. 127–147. [Google Scholar]
- Musselman, C.; Kutateladze, T. PHD Fingers: Epigenetic Effectors and Potential Drug Targets. Mol. Interv. 2009, 9, 314–323. [Google Scholar] [CrossRef]
- Huang, Q.; Zhang, L.; Wang, Y.; Zhang, C.; Zhou, S.; Yang, G.; Li, Z.; Gao, X.; Chen, Z.; Zhang, Z. Depletion of PHF14, a novel histone-binding protein gene, causes neonatal lethality in mice due to respiratory failure. Acta Biochim. Biophys. Sin. 2013, 45, 622–633. [Google Scholar] [CrossRef] [Green Version]
- Kitagawa, M.; Takebe, A.; Ono, Y.; Imai, T.; Nakao, K.; Nishikawa, S.-I.; Era, T. Phf14, a Novel Regulator of Mesenchyme Growth via Platelet-derived Growth Factor (PDGF) Receptor-α. J. Biol. Chem. 2012, 287, 27983–27996. [Google Scholar] [CrossRef] [Green Version]
- Hassoun, J.; Gambarelli, D.; Grisoli, F.; Pellet, W.; Salamon, G.; Pellissier, J.F.; Toga, M. Central neurocytoma. Acta Neuropathol. 1982, 56, 151–156. [Google Scholar] [CrossRef]
- Rades, D.; Fehlauer, F. Treatment options for central neurocytoma. Neurology 2002, 59, 1268–1270. [Google Scholar] [CrossRef]
- Gaetano, C.; Manni, I.; Bossi, G.; Piaggio, G.; Soddu, S.; Farina, A.; Helman, L.; Sacchi, A. Retinoic acid and camp differentially regulate human chromogranin a promoter activity during differentiation of neuroblastoma cells. Eur. J. Cancer 1995, 31, 447–452. [Google Scholar] [CrossRef]
- Zhang, D.; Babayan, L.; Ho, H.; Heaney, A.P. Chromogranin A regulates neuroblastoma proliferation and phenotype. Biol. Open 2019, 8, bio036566. [Google Scholar] [CrossRef] [Green Version]
- Andrae, J.; Gallini, R.; Betsholtz, C. Role of platelet-derived growth factors in physiology and medicine. Genes Dev. 2008, 22, 1276–1312. [Google Scholar] [CrossRef] [Green Version]
- Heldin, C.-H. Targeting the PDGF signaling pathway in tumor treatment. Cell Commun. Signal. 2013, 11, 97. [Google Scholar] [CrossRef]
- Goldman, J.M.; Melo, J.V. Targeting the BCR-ABL Tyrosine Kinase in Chronic Myeloid Leukemia. N. Engl. J. Med. 2001, 344, 1084–1086. [Google Scholar] [CrossRef] [PubMed]
- Demetri, G.D.; van Oosterom, A.T.; Garrett, C.R.; Blackstein, M.E.; Shah, M.H.; Verweij, J.; McArthur, G.; Judson, I.R.; Heinrich, M.C.; Morgan, J.A.; et al. Efficacy and safety of sunitinib in patients with advanced gastrointestinal stromal tumour after failure of imatinib: A randomised controlled trial. Lancet 2006, 368, 1329–1338. [Google Scholar] [CrossRef]
- Motzer, R.J.; Hutson, T.E.; Cella, D.; Reeves, J.; Hawkins, R.; Guo, J.; Nathan, P.; Staehler, M.; De Souza, P.; Merchan, J.R.; et al. Pazopanib versus Sunitinib in Metastatic Renal-Cell Carcinoma. N. Engl. J. Med. 2013, 369, 722–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, E.; Dahan, L.; Raoul, J.-L.; Bang, Y.-J.; Borbath, I.; Lombard-Bohas, C.; Valle, J.; Metrakos, P.; Smith, D.; Vinik, A.; et al. Sunitinib Malate for the Treatment of Pancreatic Neuroendocrine Tumors. N. Engl. J. Med. 2011, 364, 501–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paek, S.H.; Shin, H.Y.; Kim, J.W.; Park, S.-H.; Son, J.H.; Kim, D.G. Primary Culture of Central Neurocytoma: A Case Report. J. Korean Med. Sci. 2010, 25, 798–803. [Google Scholar] [CrossRef] [Green Version]
- Westphal, M.; Meissner, H.; Matschke, J.; Herrmann, H.-D. Tissue culture of human neurocytomas induces the expression of glial fibrilary acidic protein. J. Neurocytol. 1998, 27, 805–816. [Google Scholar] [CrossRef]
- Englot, D.J.; Chang, E.F.; Gupta, N. Neuronal Tumors. In Pediatric CNS Tumors; Gupta, N., Banerjee, A., Haas-Kogan, D.A., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 171–186. [Google Scholar]
- Li, Y.; Ye, X.-F.; Qian, G.; Yin, Y.; Pan, Q.-G. Pathologic features and clinical outcome of central neurocytoma: Analysis of 15 cases. Chin. J. Cancer Res. 2012, 24, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Mut, M.; Güler-Tezel, G.; Lopes, M.B.S.; Bilginer, B.; Ziyal, I.; E Ozcan, O. Challenging diagnosis: Oligodendroglioma versus extraventricular neurocytoma. Clin. Neuropathol. 2005, 24, 225–229. [Google Scholar]
- Beltran, H.; Eng, K.; Mosquera, J.M.; Sigaras, A.; Romanel, A.; Rennert, H.; Kossai, M.; Pauli, C.; Faltas, B.; Fontugne, J.; et al. Whole-Exome Sequencing of Metastatic Cancer and Biomarkers of Treatment Response. JAMA Oncol. 2015, 1, 466–474. [Google Scholar] [CrossRef]
- Baker, L.A.; Allis, C.D.; Wang, G.G. PHD fingers in human diseases: Disorders arising from misinterpreting epigenetic marks. Mutat. Res. Mol. Mech. Mutagen. 2008, 647, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Akazawa, T.; Yasui, K.; Gen, Y.; Yamada, N.; Tomie, A.; Dohi, O.; Mitsuyoshi, H.; Yagi, N.; Itoh, Y.; Naito, Y.; et al. Aberrant expression of the PHF14 gene in biliary tract cancer cells. Oncol. Lett. 2013, 5, 1849–1853. [Google Scholar] [CrossRef]
- Ivanov, I.; Lo, K.C.; Hawthorn, L.; Cowell, J.K.; Ionov, Y. Identifying candidate colon cancer tumor suppressor genes using inhibition of nonsense-mediated mRNA decay in colon cancer cells. Oncogene 2006, 26, 2873–2884. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Huang, Q.; Lou, J.; Zou, L.; Wang, Y.; Zhang, P.; Yang, G.; Zhang, J.; Yu, L.; Yan, D.; et al. A novel PHD-finger protein 14/KIF4A complex overexpressed in lung cancer is involved in cell mitosis regulation and tumorigenesis. Oncotarget 2017, 8, 19684–19698. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Gremeaux, L.; Fu, Q.; Liekens, D.; Van Laere, S.; Vankelecom, H. Pituitary Progenitor Cells Tracked Down by Side Population Dissection. Stem Cells 2009, 27, 1182–1195. [Google Scholar] [CrossRef]
- Zhang, D.; Li, J.; Gao, J.; Huang, C. c-Jun/AP-1 pathway-mediated cyclin D1 expression participates in low dose arsenite-induced transformation in mouse epidermal JB6 Cl41 cells. Toxicol. Appl. Pharmacol. 2009, 235, 18–24. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Yong, W.H.; Movassaghi, M.; Rodriguez, F.J.; Yang, I.; McKeever, P.; Qian, J.; Li, J.Y.; Mao, Q.; Newell, K.L.; et al. Whole Exome Sequencing Identifies PHF14 Mutations in Neurocytoma and Predicts Responsivity to the PDGFR Inhibitor Sunitinib. Biomedicines 2022, 10, 2842. https://doi.org/10.3390/biomedicines10112842
Zhang D, Yong WH, Movassaghi M, Rodriguez FJ, Yang I, McKeever P, Qian J, Li JY, Mao Q, Newell KL, et al. Whole Exome Sequencing Identifies PHF14 Mutations in Neurocytoma and Predicts Responsivity to the PDGFR Inhibitor Sunitinib. Biomedicines. 2022; 10(11):2842. https://doi.org/10.3390/biomedicines10112842
Chicago/Turabian StyleZhang, Dongyun, William H. Yong, Masoud Movassaghi, Fausto J. Rodriguez, Issac Yang, Paul McKeever, Jiang Qian, Jian Yi Li, Qinwen Mao, Kathy L. Newell, and et al. 2022. "Whole Exome Sequencing Identifies PHF14 Mutations in Neurocytoma and Predicts Responsivity to the PDGFR Inhibitor Sunitinib" Biomedicines 10, no. 11: 2842. https://doi.org/10.3390/biomedicines10112842