
Some Aspects of the Physiology of the Nyctotherus velox, a Commensal Ciliated Protozoon Taken from the Hindgut of the Tropical Millipede Archispirostreptus gigas
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Ciliates
2.2. Identification of Nyctotherus Species
2.3. Cultivation of Nyctotherus In Vitro
2.4. Experiments on the Growth Requirements of the Nyctotherus
2.4.1. Experiment 1
2.4.2. Experiment 2
2.4.3. Experiment 3
2.5. Counting of Ciliates and Total Bacteria
2.6. Morphometry of Trophozoites and Cysts
2.7. Measurement of Fermentation Activity
2.8. Measurement of Hydrolytic Enzymatic Activities
2.8.1. Harvesting the Ciliates
2.8.2. Preparation of Crude Protein Extracts for the Measurement of Hydrolytic Activities
2.8.3. The Measurement of Protein Concentration
2.8.4. The Measurement of Hydrolytic Activities
2.9. Statistical Analysis
3. Results
3.1. Effects of Temperatures and Soluble Nutrients
3.2. Effects of Plant Polysaccharide Substrates
3.3. Growth in Aerobic Conditions
3.4. Fermentation Patterns
3.5. Enzyme Hydrolytic Activities
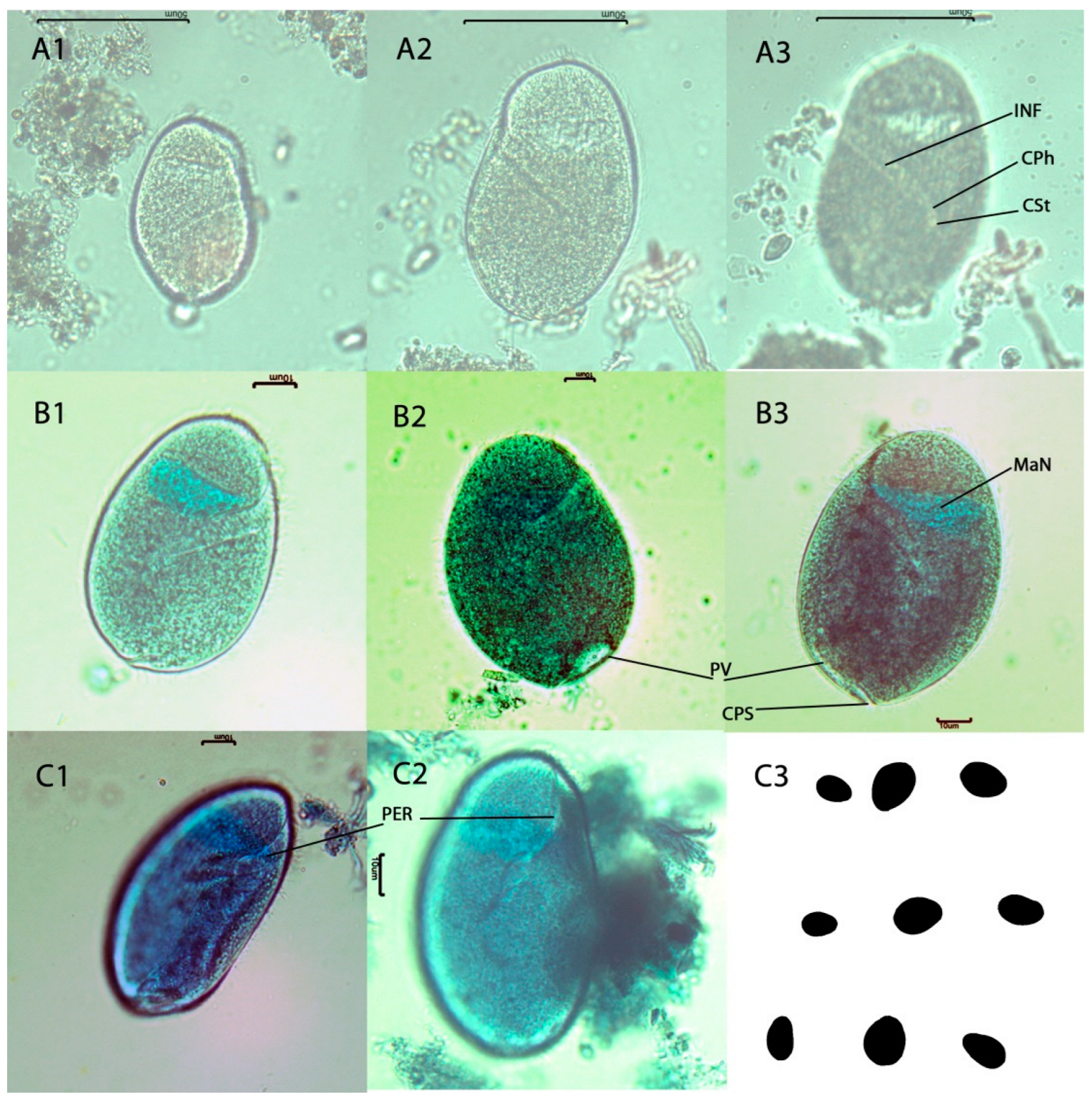
3.6. Molecular and Morphological Description of Ciliates
- Taxonomic summary
- Phylum Ciliophora Doflein, 1901
- Subphylum Intramacronucleata Lynn, 1996
- Class Armophorea Lynn, 2004
- Order Clevelandellida de Puytorac and Grain, 1976
- Family Nyctotheridae Amaro, 1972
- Genus Nyctotherus Leidy, 1849
- Type locality: Hindgut of the millipede Archispirostreptus gigas.
- Type host: Archispirostreptus gigas Peters, 1868
- Country of the host origin: Tanzania
- Gene sequences. The nuclear 18S rDNA gene sequences were deposited in GenBank. Accession numbers are listed in the Supplementary Materials.
- Family Nyctotheridae Amaro, 1972
- Genus Nyctotherus Leidy, 1849
- Type locality: Hindgut of the millipede Archispirostreptus gigas.
- Type host: Archispirostreptus gigas Peters, 1868
- Country of the host origin: Tanzania
- Gene sequences: The nuclear 18S rDNA gene sequences were deposited in GenBank. Accession numbers are listed in the Supplementary Materials.
- Etymology: The species name is derived from the generic name of the host.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Albaret, J.L. Étude systématique et cytologique sur les ciliés hétérotriches endocommensaux (Systematic and cytological study on heterotrichous endocommensal ciliates). Mémoires Muséum Natl. D’histoire Nat. Ser. A Zool. 1975, 89, 1–114. [Google Scholar]
- Gijzen, H.J.; Broers, C.A.; Barughare, M.; Stumm, C.K. Methanogenic bacteria as endosymbionts of the ciliate Nyctotherus ovalis in the cockroach hindgut. Appl. Environ. Microbiol. 1991, 57, 1630–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gijzen, H.J.; Barugahare, M. Contribution of anaerobic protozoa and methanogens to hindgut metabolic activities of the American cockroach, Periplaneta americana. Appl. Environ. Microbiol. 1992, 58, 2565–2570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gijzen, H.J.; van der Drift, C.; Barugahare, M.; Op den Camp, H.J. Effect of Host Diet and Hindgut Microbial Composition on Cellulolytic Activity in the Hindgut of the American Cockroach, Periplaneta americana. Appl. Environ. Microbiol. 1994, 60, 1822–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Graaf, R.M.; Ricard, G.; van Alen, T.A.; Duarte, I.; Dutilh, B.E.; Burgtorf, C.; Kuiper, J.W.P.; van der Staay, G.W.M.; Tielens, A.G.M.; Huynen, M.A.; et al. The Organellar Genome and Metabolic Potential of the Hydrogen-Producing Mitochondrion of Nyctotherus ovalis. Mol. Biol. Evol. 2011, 28, 2379–2391. [Google Scholar] [CrossRef] [Green Version]
- Balch, H.E. The Cultivation of Nyctotherus ovalis and Endamoeba blattae. Science 1967, 76, 237. [Google Scholar] [CrossRef]
- Nelson, E.C. Cultivation of Nyctotherus cordiformis. J. Parasitol. 1943, 29, 292. [Google Scholar] [CrossRef]
- Lom, J. Experiments with the cultivation of our three species of the genus Nyctotherus and of Balantidium entozoon and B. coli. Věst. Českoslov. Zool. Spol. 1956, 20, 16–61. [Google Scholar]
- Chen, L. Züchtungsversuche an parasitischen Protozoen von Periplaneta orientalis. Zeitschrift. Parasitenkd. 1933, 6, 207–219. [Google Scholar] [CrossRef]
- Byzov, B.A. Intestinal Microbiota of Millipedes. In Intestinal Microorganisms of Termites and Other Invertebrates; König, H., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 89–114. [Google Scholar] [CrossRef]
- Šustr, V.; Tajovský, K.; Semanová, S.; Chroňáková, A.; Šimek, M. The giant African millipede, Archispirostreptus gigas (Diplopoda: Spirostreptida), a model species for ecophysiological studies. Acta Soc. Zool. Bohem. 2013, 77, 145–158. [Google Scholar]
- Nardi, J.B.; Bee, C.M.; Taylor, S.J. Compartmentalization of microbial communities that inhabit the hindguts of millipedes. Arthropod Struct. Dev. 2016, 45, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.C. Role of Aerobic Microbial Populations in Cellulose Digestion by Desert Millipedes. Appl. Environ. Microbiol. 1982, 44, 281–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šustr, V.; Semanová, S.; Rost-Roszkowska, M.M.; Tajovský, K.; Sosinka, A.; Kaszuba, F. Enzymatic activities in the digestive tract of spirostreptid and spirobolid millipedes (Diplopoda: Spirostreptida and Spirobolida). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2020, 241, 110388. [Google Scholar] [CrossRef]
- Regensbogenova, M.; Kisidayova, S.; Michalowski, T.; Javorsky, P.; Moon-Van Der Staay, S.Y.; Moon-Van Der Staay, G.W.M.M.; Hackstein, J.H.P.; Mcewan, N.R.; Jouany, J.-P.; Newbold, J.C.; et al. Rapid identification of rumen protozoa by restriction analysis of amplified 18S rRNA gene. Acta Protozool. 2004, 43, 219–224. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; pp. 95–98. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony and Other Methods, Version 4; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Foissner, W. An update of “basic light and scanning electron microscopic methods for taxonomic studies of ciliated protozoa”. Int. J. Syst. Evol. Microbiol. 2014, 64, 271–292. [Google Scholar] [CrossRef] [Green Version]
- Foissner, W. Basic light and scanning electron microscopic methods for taxonomic studies of ciliated protozoa. Eur. J. Protistol. 1991, 27, 313–330. [Google Scholar] [CrossRef] [PubMed]
- Doddema, H.J.; Vogels, G.D. Improved identification of methanogenic bacteria by fluorescence microscopy. Appl. Environ. Microbiol. 1978, 36, 752–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.G.; Coleman, G.S. The Rumen Protozoa; Springer: New York, NY, USA, 1992. [Google Scholar] [CrossRef]
- Kišidayová, S.; Váradyová, Z.; Zeleňák, I.; Siroka, P. Methanogenesis in rumen ciliate cultures of Entodinium caudatum and Epidinium ecaudatum after long-term cultivation in a chemically defined medium. Folia Microbiol. 2000, 45, 269–274. [Google Scholar] [CrossRef]
- Hino, T.; Kametaka, M.; Kandatsu, M. The cultivation of rumen oligotrich protozoa. III. White clover factors which stimulate the growth of Entodinia. J. Gen. Appl. Microbiol. 1973, 19, 397–413. [Google Scholar] [CrossRef] [Green Version]
- Coleman, G.S. Rumen entodiniomorphid protozoa. In Methods of Cultivating Parasites In Vitro; Taylor, A.E.R., Baker, J.R., Eds.; Academic Press Inc.: London, UK, 1978; pp. 39–54. [Google Scholar]
- Horáková, K. Mikroskopické metódy (Microscopic methods). In Mikrobiologické Laboratórne Metódy (Microbiological Laboratory Methods); Betina, V., Barátová, H., Fargašová, A., Frank, V., Horáková, K., Šturdík, E., Eds.; Alfa: Bratislava, Slovakia, 1988; pp. 173–206. [Google Scholar]
- Grishagin, I.V. Automatic cell counting with ImageJ. Anal. Biochem. 2015, 473, 63–65. [Google Scholar] [CrossRef] [PubMed]
- Abramoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Kišidayová, S.; Váradyová, Z. Effect of insulin on in vitro fermentation activity of microrganism community of rumen ciliate culture. Cell Biol. Int. 2005, 29, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Cottyn, B.G.; Boucque, C.V. Rapid method for the gas-chromatographic determination of volatile fatty acids in rumen fluid. J. Agric. Food Chem. 1968, 16, 105–107. [Google Scholar] [CrossRef]
- Chaney, A.L.; Marbach, E.P. Modified Reagents for Determination of Urea and Ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef]
- Mellenberger, R.W.; Satter, L.D.; Millett, M.A.; Baker, A.J. An in vitro Technique for Estimating Digestibility of Treated and Untreated Wood. J. Anim. Sci. 1970, 30, 1005–1011. [Google Scholar] [CrossRef] [Green Version]
- Nevel, C.J.; Demeyer, D.I. Stoichiometry of carbohydrate fermentation and microbial growth efficiency in a continous culture of mixed rumen bacteria. Eur. J. Appl. Microbiol. Biotechnol. 1979, 7, 111–120. [Google Scholar] [CrossRef]
- Ungerfeld, E.M. Shifts in metabolic hydrogen sinks in the methanogenesis-inhibited ruminal fermentation: A meta-analysis. Front. Microbiol. 2015, 6, 37. [Google Scholar] [CrossRef]
- Čársky, J. Príprava presných, tlmivých a izotonických roztokovo (Preparation of precise, buffered and isotonic solutions). In Biochemické Laboratórne Metódy (Biochemical Laboratory Methods); Ferenčík, M., Škárka, B., Eds.; ALFA-VTEL: Bratislava, Slovakia, 1981; pp. 39–71. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bailey, M.J.; Biely, P.; Poutanen, K. Interlaboratory testing of methods for assay of xylanase activity. J. Biotechnol. 1992, 23, 257–270. [Google Scholar] [CrossRef]
- Béra-Maillet, C.; Devillard, E.; Cezette, M.; Jouany, J.-P.; Forano, E. Xylanases and carboxymethylcellulases of the rumen protozoa Polyplastron multivesiculatum, Eudiplodinium maggii, and Entodinium sp. FEMS Microbiol. Lett. 2005, 244, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.L.; Blum, R.; Glennon, W.E.; Burton, A.L. Measurement of carboxymethylcellulase activity. Anal. Biochem. 1960, 1, 127–132. [Google Scholar] [CrossRef]
- Small, E.B.; Marszalek, D.S.; Antipa, G.A. A Survey of Ciliate Surface Patterns and Organelles as Revealed with Scanning Electron Microscopy. Trans. Am. Microsc. Soc. 1971, 90, 283. [Google Scholar] [CrossRef]
- Imai, S.; Katsuno, M.; Tsunoda, K. Scanning Electron Microscopy of Rumen Ciliates in Cattle. Zool. Mag. 1977, 86, 194–207. [Google Scholar]
- Wang, Q.; Zhao, X. A three-dimensional phase diagram of growth-induced surface instabilities. Sci. Rep. 2015, 5, 8887. [Google Scholar] [CrossRef] [Green Version]
- Lynn, D.H. The implications of recent descriptions of kinetid structure to the systematics of the ciliated protists. Protoplasma 1991, 164, 123–142. [Google Scholar] [CrossRef]
- De Beaurepaire Aragão, B. Noticia sobre o Nyctotherus cordiformis Stein. Mem. Inst. Oswaldo Cruz. 1912, 4, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Pinto, C. Anatomia e biologia dos Nyctotherus do batrachios do Brasil. Bol. Biol. 1926, 3, 45–48. [Google Scholar]
- Nelson, E.C. Alcohol–preserved tissue–cultivation medium: Methods of preparation and use and results obtained in the cultivation of Nyctotherus cordiformis. Am. J. Epidemiol. 1943, 38, 185–192. [Google Scholar] [CrossRef]
- Lucas, C.L.T. A Study of Excystation in Nyctotherus ovalis: With Notes on Other Intestinal Protozoa of the Cockroach. J. Parasitol. 1928, 14, 161. [Google Scholar] [CrossRef]
- Kudo, R.R. Studies on Nyctotherus ovalis Leidy, with special reference to its nuclear structure. Arch. Protistenkd. 1936, 87, 10–42. [Google Scholar]
- Suzuki, J.; Kobayashi, S.; Yoshida, N.; Azuma, Y.; Kobayashi-Ogata, N.; Kartikasari, D.P.; Yanagawa, Y.; Iwata, S. Phylogenetic position of Nyctotherus teleacus isolated from a tortoise (Astrochelys radiata) and its electron microscopic features. J. Vet. Med. Sci. 2020, 82, 699–703. [Google Scholar] [CrossRef] [Green Version]
- Hoyte, H.M.D. The protozoa occurring in the hind-gut of cockroaches. I. Responses to changes in environment. Parasitology 1961, 51, 415–436. [Google Scholar] [CrossRef]
- Lom, J. Polysacharidové reservy nálevníků rodu Balantidium a Nyctotherus. Českoslov. Biol. 1955, 4, 394–409. [Google Scholar]
- Dutta, G.P. The Cytoplasmic Inclusions of Nyctotherus macropharyngeus: Histochemical Studies. J. Cell Sci. 1958, s3-99, 517–521. [Google Scholar] [CrossRef]
- Hoyte, H.M.D. The protozoa occurring in the hind-gut of cockroaches. II. Morphology of Nyctotherus ovalis. Parasitology 1961, 51, 437–463. [Google Scholar] [CrossRef]
- Nakai, Y.; Imai, S. Cytochemical Identification of Reserve Polysaccharides in Rumen Ciliates by Microspectrophotometry. Jpn. J. Parasitol. 1989, 38, 216–220. [Google Scholar]
- Brust, H.; Orzechowski, S.; Fettke, J. Starch and Glycogen Analyses: Methods and Techniques. Biomolecules 2020, 10, 1020. [Google Scholar] [CrossRef]
- Wakita, M.; Hoshino, S. Physicochemical properties of a reserve polysaccharide from sheep rumen ciliates genus Entodinium. Comp. Biochem. Physiol. Part B Comp. Biochem. 1980, 65, 571–574. [Google Scholar] [CrossRef]
- Wani, A.A.; Singh, P.; Shah, M.A.; Schweiggert-Weisz, U.; Gul, K.; Wani, I.A. Rice Starch Diversity: Effects on Structural, Morphological, Thermal, and Physicochemical Properties-A Review. Compr. Rev. Food Sci. Food Saf. 2012, 11, 417–436. [Google Scholar] [CrossRef]
- Zhao, L.; Pan, T.; Guo, D.; Wei, C. A simple and rapid method for preparing the whole section of starchy seed to investigate the morphology and distribution of starch in different regions of seed. Plant Methods 2018, 14, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horváthová, T.; Šustr, V.; Chroňáková, A.; Semanová, S.; Lang, K.; Dietrich, C.; Hubáček, T.; Ardestani, M.M.; Lara, A.C.; Brune, A.; et al. Methanogenesis in the Digestive Tracts of the Tropical Millipedes Archispirostreptus gigas (Diplopoda, Spirostreptidae) and Epibolus pulchripes (Diplopoda, Pachybolidae). Appl. Environ. Microbiol. 2021, 87, e0061421. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.E.; Williams, A.G.; Lloyd, D. Oxygen consumption by ruminal microorganisms: Protozoal and bacterial contributions. Appl. Environ. Microbiol. 1989, 55, 2583–2587. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.E.; Mcintyre, P.S.; Saleh, M.; Williams, A.G.; Lloyd, D. The influence of ruminal concentrations of O2 and CO2 on fermentative metabolism of the rumen entodiniomorphid ciliate Eudiplodinium maggii. Curr. Microbiol. 1991, 23, 245–251. [Google Scholar] [CrossRef]
- Park, T.; Yu, Z. Aerobic cultivation of anaerobic rumen protozoa, Entodinium caudatum and Epidinium caudatum. J. Microbiol. Methods 2018, 152, 186–193. [Google Scholar] [CrossRef]
- Fievez, V.; Piattoni, F.; Mbanzamihigo, L.; Demeyer, D. Reductive Acetogenesis in the Hindgut and Attempts to its Induction in the Rumen—A Review. J. Appl. Anim. Res. 1999, 16, 1–22. [Google Scholar] [CrossRef]
- Laho, T.; Váradyová, Z.; Mihaliková, K.; Kišidayová, S. Fermentation Capacity of Fecal Microbial Inocula of Przewalski Horse, Kulan, and Chapman Zebra and Polysaccharide Hydrolytic Activities of Fecal Microbial Constituents (Ciliates and Bacteria) of Kulan and Chapman Zebra. J. Equine Vet. Sci. 2013, 33, 143–149. [Google Scholar] [CrossRef]
- Kišidayová, S.; Váradyová, Z.; Pristaš, P.; Piknová, M.; Nigutová, K.; Petrželková, K.J.; Profousová, I.; Schovancová, K.; Kamler, J.; Modrý, D. Effects of high- and low-fiber diets on fecal fermentation and fecal microbial populations of captive chimpanzees. Am. J. Primatol. 2009, 71, 548–557. [Google Scholar] [CrossRef]
- Cieslak, A.; Váradyová, Z.; Kišidayová, S.; Szumacher-Strabel, M. The effects of linoleic acid on the fermentation parameters, population density, and fatty-acid profile of two rumen ciliate cultures, Entodinium caudatum and Diploplastron affine. Acta Protozool. 2009, 48, 51–61. [Google Scholar]
- Ivan, M.; Neill, L.; Forster, R.; Alimon, R.; Rode, L.M.; Entz, T. Effects of Isotricha, Dasytricha, Entodinium, and Total Fauna on Ruminal Fermentation and Duodenal Flow in Wethers Fed Different Diets. J. Dairy Sci. 2000, 83, 776–787. [Google Scholar] [CrossRef] [PubMed]
- Boxma, B.; de Graaf, R.M.; van der Staay, G.W.M.; van Alen, T.A.; Ricard, G.; Gabaldón, T.; Van Hoek, A.H.A.M.; Der Staay, S.Y.M.-V.; Koopman, W.J.H.; Van Hellemond, J.J.; et al. An anaerobic mitochondrion that produces hydrogen. Nature 2005, 434, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Šustr, V.; Chroňáková, A.; Semanová, S.; Tajovský, K.; Šimek, M. Methane production and methanogenic Archaea in the digestive tracts of millipedes (Diplopoda). PLoS ONE 2014, 9, e102659. [Google Scholar] [CrossRef] [Green Version]
- Finlay, B.J.; Esteban, G.; Clarke, K.J.; Williams, A.G.; Embley, T.M.; Hirt, R.P. Some rumen ciliates have endosymbiotic methanogens. FEMS Microbiol. Lett. 1994, 117, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Lewis, W.H.; Sendra, K.M.; Embley, T.M.; Esteban, G.F. Morphology and Phylogeny of a New Species of Anaerobic Ciliate, Trimyema finlayi n. sp., with Endosymbiotic Methanogens. Front. Microbiol. 2018, 9, 140. [Google Scholar] [CrossRef]
- Vogels, G.D.; Hoppe, W.F.; Stumm, C.K. Association of methanogenic bacteria with rumen ciliates. Appl. Environ. Microbiol. 1980, 40, 608–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tóthová, T.; Piknová, M.; Kišidayová, S.; Javorský, P.; Pristaš, P. Distinctive archaebacterial species associated with anaerobic rumen protozoan Entodinium caudatum. Folia Microbiol. 2008, 53, 259–262. [Google Scholar] [CrossRef]
- Stumm, C.K.; Zwart, K.B. Symbiosis of protozoa with hydrogen-utilizing methanogens. Microbiol. Sci. 1986, 3, 100–105. [Google Scholar] [PubMed]
- Goosen, N.K.; Horemans, A.M.C.; Hillebrand, S.J.W.; Stumm, C.K.; Vogels, G.D. Cultivation of the sapropelic ciliate Plagiopyla nasuta Stein and isolation of the endosymbiont Methanobacterium formicicum. Arch. Microbiol. 1988, 150, 165–170. [Google Scholar] [CrossRef]
- Regensbogenova, M.; Michalowski, T.; Newbold, C.; McEwan, N.; Javorsky, P.; Kišidayová, S.; Hackstein, J.; Pristaš, P. A re-appraisal of the diversity of the methanogens associated with the rumen ciliates. FEMS Microbiol. Lett. 2004, 238, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Nunez, F.S.; Crawford, C.S. Digestive enzymes of the desert millipede Orthoporus ornatus (Girard) (Diplopoda: Spirostreptidae). Comp. Biochem. Physiol. Part A Physiol. 1976, 55, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Profousová, I.; Mihaliková, K.; Laho, T.; Váradyová, Z.; Petrželková, K.J.; Modrý, D.; Kišidayová, S. The ciliate, Troglodytella abrassarti, contributes to polysaccharide hydrolytic activities in the chimpanzee colon. Folia Microbiol. 2011, 56, 339–343. [Google Scholar] [CrossRef]
- Kišidayová, S.; Pristaš, P.; Zimovčáková, M.; Blanár Wencelová, M.; Homol’ová, L.; Mihaliková, K.; Čobanová, K.; Grešáková, Ľ.; Váradyová, Z. The effects of high dose of two manganese supplements (organic and inorganic) on the rumen microbial ecosystem. PLoS ONE 2018, 13, e0191158. [Google Scholar] [CrossRef] [Green Version]
- González, G.; Murphy, C.M.; Belén, J. Direct and Indirect Effects of Millipedes on the Decay of Litter of Varying Lignin Content. In Tropical Forests; Sudarshana, P., Nageswara-Rao, M., Soneji, J.R., Eds.; InTech: London, UK, 2012; pp. 37–50. [Google Scholar] [CrossRef] [Green Version]
- Demeyer, D.I.; De Graeve, K. Differences in Stoichiometry Between Rumen and Hindgut Fermentation. Adv. Anim. Physiol. Anim. Nutr. 1991, 64, 50–61. [Google Scholar]
- Sardar, P.; Šustr, V.; Chroňáková, A.; Lorenc, F. Metatranscriptomic holobiont analysis of carbohydrate-active enzymes in the millipede Telodeinopus aoutii (Diplopoda, Spirostreptida). Front. Ecol. Evol. 2022, 10, 873. [Google Scholar] [CrossRef]
- Hogan, M.E.; Slaytor, M.; O’Brien, R.W. Transport of volatile fatty acids across the hindgut of the cockroach Panesthia cribrata Saussure and the termite, Mastotermes darwiniensis Froggatt. J. Insect Physiol. 1985, 31, 587–591. [Google Scholar] [CrossRef]
- Leidy, J. Some Observations on Nematoidea Imperfecta, and Descriptions of Three Parasitic Infusoriae. Trans. Am. Philos. Soc. 1853, 10, 241–246. [Google Scholar] [CrossRef]
- Leidy, J. Nyctotherus, a new genus of Polygastrica, allied to Plescoma. Proc. Acad. Nat. Sci. USA 1849, 4, 233. [Google Scholar]
- Savin, M.B. Nyctotherus velox and Endolimax sp. with Remarks on Other Prozozoan Parasites Found in the Millipede, Spirobolus marginatus. Master’s Thesis, University of Pennsylvania, Philadelphia, PA, USA, 1931. [Google Scholar]
- van Hoek, A.H.; van Alen, T.A.; Sprakel, V.S.; Hackstein, J.H.; Vogels, G.D. Evolution of anaerobic ciliates from the gastrointestinal tract: Phylogenetic analysis of the ribosomal repeat from Nyctotherus ovalis and its relatives. Mol. Biol. Evol. 1998, 15, 1195–1206. [Google Scholar] [CrossRef]
- Lynn, D.H.; Wright, A.-D.G. Biodiversity and Molecular Phylogeny of Australian Clevelandella Species (Class Armophorea, Order Clevelandellida, Family Clevelandellidae), Intestinal Endosymbiotic Ciliates in the Wood-Feeding Roach Panesthia cribrata Saussure, 1864. J. Eukaryot. Microbiol. 2013, 60, 335–341. [Google Scholar] [CrossRef]
- Li, M.; Hu, G.; Li, C.; Zhao, W.; Zou, H.; Li, W.; Wu, S.-G.; Wang, G.-T.; Ponce-Gordo, F. Morphological and molecular characterization of a new ciliate Nyctotheroides grimi n. sp. (Armophorea, Clevelandellida) from Chinese frogs. Acta Trop. 2020, 208, 105531. [Google Scholar] [CrossRef] [PubMed]
- Pecina, L.; Vďačný, P. Two New Endozoic Ciliates, Clevelandella lynni sp. n. and Nyctotherus galerus sp. n., Isolated from the Hindgut of the Wood-feeding Cockroach Panesthia angustipennis (Illiger, 1801). J. Eukaryot. Microbiol. 2020, 67, 436–449. [Google Scholar] [CrossRef]
- Paiva, T.D.S.; Borges, B.D.N.; Silva-Neto, I.D.D. Phylogenetic study of Class Armophorea (Alveolata, Ciliophora) based on 18S-rDNA data. Genet. Mol. Biol. 2013, 36, 571–585. [Google Scholar] [CrossRef]
- Pecina, L.; Vďačný, P. Morphological versus molecular delimitation of ciliate species: A case study of the family Clevelandellidae (Protista, Ciliophora, Armophorea). Eur. J. Taxon. 2020, 697, 1–46. [Google Scholar] [CrossRef]
- Affa’a, F.M.; Hickey, D.A.; Strüder-Kypke, M.; Lynn, D.H. Phylogenetic position of species in the genera Anoplophrya, Plagiotoma, and Nyctotheroides (Phylum ciliophora), endosymbiotic ciliates of annelids and anurans. J. Eukaryot. Microbiol. 2004, 51, 301–306. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rice Starch | CMC | Celulose20 | Xylan | Inulin | In Situ | NoPOS | |
|---|---|---|---|---|---|---|---|
| Trophozoites | |||||||
| Number, n/mL | 690 ± 240 * | 490 ± 160 | 500 ± 300 | 360 ± 140 | 390 ± 280 | ND | 450 ± 70 |
| Length, µm | 68 ± 9.2 * | 51 ± 5.1 *** | 59 ± 9.4 *** | 62 ± 7.1 *** | 61 ± 12.9 *** | 75 ± 9.2 | 55 ± 6.0 *** |
| Width, µm | 43 ± 7.9 ** | 33 ± 4.0 *** | 42 ± 9.0 ** | 40 ± 5.4 *** | 38 ± 12.0 *** | 50 ± 8.1 | 36 ± 4.4 *** |
| L/W | 1.6 ± 0.2 | 1.5 ± 0.2 | 1.5 ± 0.2 | 1.5 ± 0.9 | 1.7 ± 0.2 | 1.5 ± 0.2 | 1.6 ± 0.2 |
| Cysts | |||||||
| Number, n/mL | 160 ± 100 | 200 ± 150 | 130 ± 100 | 140 ± 90 | 60 ± 40 * | ND | 290 ± 90 |
| Length, µm | 30 ± 1.9 | 27 ± 3.9 | 30 ± 3.0 | 28 ± 1.8 | 33 ± 3.9 | - | 31 ± 0 |
| Width, µm | 24 ± 1.6 | 21 ± 1.2 | 24 ± 2.1 | 22 ± 2.0 | 24 ± 2.0 | - | 23 ± 0 |
| L/W | 1.3 ± 0 | 1.3 ± 0.1 | 1.2 ± 0 | 1.3 ± 0.1 | 1.4 ± 0.1 | - | 1.4 ± 0 |
| Bacteria (n/mL, Ln) | 22.175 ± 0.2 | 20.685 ± 0.5 * | 22.173 ± 0.5 | 21.273 ± 0.3 | 22.387 ± 0.3 | - | - |
| Item | Xylan | CMC | CC | Rice Starch | Inulin | NoPOS | SE | p-Value | |
|---|---|---|---|---|---|---|---|---|---|
| IVDMD | g/kg DMsubst | 48.10 c,d | 59.27 | 50.59 a,b | 92.46 b,d | 88.30 a,c | - | 5.211 | 0.0019 |
| Gas | mL/g DMsubst | 75 a,d,g | 16 b,e | 4 c,f,g | 174 a,b,c | 150 d,e,f | - | 16.156 | 0.0001 |
| Methane | % (vol/vol) | 17.89 a,b,c,d,e | 4.11 a | 8.77 b | 5.29 c | 10.19 d | 3.85 e | 1.182 | 0.0001 |
| Ammonia | mg/L | 20.94 a,b,c | 144.00 a,d,g | 148.63 b,e,h | 32.88 d,e,f | 30.78 g,h,i | 143.50 c,f,i | 12.627 | 0.0001 |
| SCFA | mmol/L | 22.89 a | 3.47 | 5.43 | 45.45 b | 40.63 c | 5.60 a,b,c | 3.581 | 0.0001 |
| Acetate | mol% | 62.69 a | 83.89 | 74.78 b | 65.72 c | 67.91 d | 82.36 a,b,c,d | 1.719 | 0.0001 |
| Propionate | mol% | 29.72 a | 1.38 | 14.14 b | 21.99 c | 23.39 d | 3.50 a,b,c,d | 2.220 | 0.0001 |
| nButyrate | mol% | 6.13 a | 0.52 | 1.72 | 9.09 b | 5.83 c | 1.51 a,b,c | 0.482 | 0.0001 |
| iButyrate | mol% | 0.62 a | 5.50 | 3.92 | 0.23 b | 0.40 c | 5.22 a,b,c | 0.649 | 0.0001 |
| nValerate | mol% | 0.52 a | 3.27 b | 2.22 | 2.67 | 2.00 | 2.10 a,b | 0.185 | 0.0001 |
| iValerate | mol% | 0.33 a | 5.36 | 3.16 b | 0.28 c | 0.47 d | 5.26 a,b,c,d | 0.474 | 0.0001 |
| 2H-recovery | % | 80.13 a | 28.23 | 47.91 b | 48.02 c | 53.93 d | 30.16 a,b,c,d | 3.787 | 0.0001 |
| Trophozoites number, T | n/mL | 608 | 823 | 650 | 115 | 800 | 558 | 70.985 | 0.0708 |
| Cysts number, C | n/mL | 108 | 243 b | 160 | 105 a,b | 175 | 263 a | 14.674 | 0.0035 |
| Cyst proportion | % | 12.05 | 22.75 b | 19.67 | 63.42 a,b | 29.98 | 36.06 a | 5.24 | 0.0035 |
| Enzyme | Catalytic Activity | Specific Catalytic Activity | ||
|---|---|---|---|---|
| nkat/L ± SD | n | nkat/g Protein ± SD | n | |
| Amylase | 59 830 ± 71 490 | 2 | 300 ± 120 | 2 |
| CM-cellulase | 14 900 ± 21 830 | 6 | 190 ± 110 | 6 |
| Xylanase | 91 930 ± 72 440 | 4 | 290 ± 100 | 4 |
| Inulinase | 87 830 ± 110 300 | 2 | 170 ± 220 | 2 |
| Ciliate Area (µm2) | Ciliate Lenght (L) (µm) | Ciliate Width (W) (µm) | L/W | MaN Lenght (µm) | MaN Width (µm) | Infund. Angle (°) | Infund. Lenght (µm) | |
|---|---|---|---|---|---|---|---|---|
| Arith. mean | 3270 | 78 | 54 | 1.5 | 30 | 10 | 125 | 28 |
| SD | 970 | 10 | 11 | 0.14 | 4.5 | 2.8 | 2.2 | 3.9 |
| Min | 1950 | 61 | 42 | 1.2 | 23 | 8 | 121 | 21 |
| Median | 3080 | 81 | 50 | 1.5 | 32 | 9 | 126 | 28 |
| Max | 4590 | 91 | 70 | 1.7 | 36 | 16 | 126 | 33 |
| n | 9 | 9 | 9 | 9 | 7 | 7 | 4 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kišidayová, S.; Scholcová, N.; Mihaliková, K.; Váradyová, Z.; Pristaš, P.; Weisskopf, S.; Chrudimský, T.; Chroňáková, A.; Šimek, M.; Šustr, V. Some Aspects of the Physiology of the Nyctotherus velox, a Commensal Ciliated Protozoon Taken from the Hindgut of the Tropical Millipede Archispirostreptus gigas. Life 2023, 13, 1110. https://doi.org/10.3390/life13051110
Kišidayová S, Scholcová N, Mihaliková K, Váradyová Z, Pristaš P, Weisskopf S, Chrudimský T, Chroňáková A, Šimek M, Šustr V. Some Aspects of the Physiology of the Nyctotherus velox, a Commensal Ciliated Protozoon Taken from the Hindgut of the Tropical Millipede Archispirostreptus gigas. Life. 2023; 13(5):1110. https://doi.org/10.3390/life13051110
Chicago/Turabian StyleKišidayová, Svetlana, Nikola Scholcová, Katarína Mihaliková, Zora Váradyová, Peter Pristaš, Stanislava Weisskopf, Tomáš Chrudimský, Alica Chroňáková, Miloslav Šimek, and Vladimír Šustr. 2023. "Some Aspects of the Physiology of the Nyctotherus velox, a Commensal Ciliated Protozoon Taken from the Hindgut of the Tropical Millipede Archispirostreptus gigas" Life 13, no. 5: 1110. https://doi.org/10.3390/life13051110