
Rickettsia Deregulates Genes Coding for the Neurotoxic Cell Response Pathways in Cerebrocortical Neurons In Vitro
 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Rickettsial Culture and Purification
2.2. Infection of Primary Neurons and Viability of Host Cells
2.3. Immunofluorescence Assay (IFA)
2.4. RNA Extraction
2.5. Gene Expression Profiling Using Real-Time PCR
2.6. Pathway Enrichment Analysis (PEA)
3. Results
3.1. Neuronal Damage Induced by R. akari and R. slovaca
3.2. Rickettsiae Induce Deregulation of Gene Expression in Primary Cerebrocortical Neurons
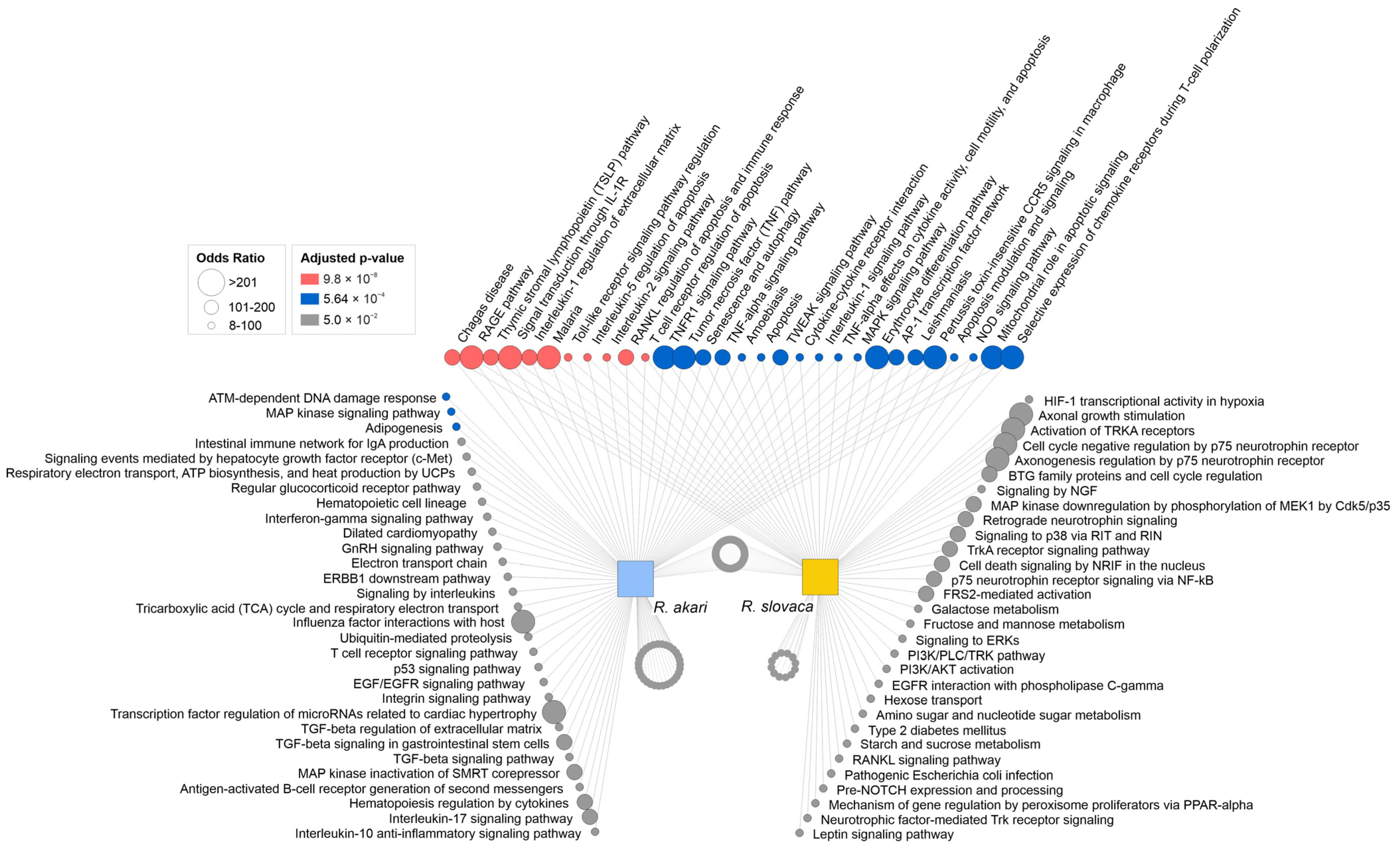
3.3. Signaling Pathways Involved in Response to Rickettsial Infection in Primary Cerebrocortical Neurons
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Venkatesan, A.; Tunkel, A.R.; Bloch, K.C.; Lauring, A.S.; Sejvar, J.; Bitnun, A.; Stahl, J.P.; Mailles, A.; Drebot, M.; Rupprecht, C.E.; et al. Case definitions, diagnostic algorithms, and priorities in encephalitis: Consensus statement of the international encephalitis consortium. Clin. Infect. Dis. 2013, 57, 1114–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatib, U.; van de Beek, D.; Lees, J.A.; Brouwer, M.C. Adults with suspected central nervous system infection: A prospective study of diagnostic accuracy. J. Infect. 2017, 74, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Glaser, C.A.; Honarmand, S.; Anderson, L.J.; Schnurr, D.P.; Forghani, B.; Cossen, C.K.; Schuster, F.L.; Christie, L.J.; Tureen, J.H. Beyond viruses: Clinical profiles and etiologies associated with encephalitis. Clin. Infect. Dis. 2006, 43, 1565–1577. [Google Scholar] [CrossRef] [Green Version]
- Dumler, J.S.; Walker, D.H. Rocky Mountain spotted fever--changing ecology and persisting virulence. N. Engl. J. Med. 2005, 353, 551–553. [Google Scholar] [CrossRef]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.H.; Fishbein, D.B. Epidemiology of rickettsial diseases. Eur. J. Epidemiol. 1991, 7, 237–245. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, C.; Su, Z.; Chang, Q.; Qiu, Y.; Bei, J.; Gaitas, A.; Xiao, J.; Drelich, A.; Khanipov, K.; et al. Endothelial Exosome Plays a Functional Role during Rickettsial Infection. mBio 2021, 12, 3. [Google Scholar] [CrossRef] [PubMed]
- Drexler, N.A.; Close, R.; Yaglom, H.D.; Traeger, M.; Parker, K.; Venkat, H.; Villarroel, L.; Brislan, J.; Pastula, D.M.; Armstrong, P.A. Morbidity and Functional Outcomes Following Rocky Mountain Spotted Fever Hospitalization-Arizona, 2002–2017. Open Forum Infect. Dis. 2022, 9, ofac506. [Google Scholar] [CrossRef]
- Sekeyová, Z.; Danchenko, M.; Filipčík, P.; Fournier, P.E. Rickettsial infections of the central nervous system. PLoS Negl. Trop. Dis. 2019, 13, e0007469. [Google Scholar] [CrossRef]
- Elzein, F.E.; Aloteibi, M.; Alanazi, W.; Alsaeed, M.; Almaghaslah, M. A rickettsia infection from Saudi Arabia. Int. J. Infect. Dis. 2020, 90, 167–169. [Google Scholar] [CrossRef]
- Bradshaw, M.J.; Lalor, K.B.; Vu, N.; Pruthi, S.; Bloch, K.C. Child Neurology: Rocky Mountain spotted fever encephalitis. Neurology 2017, 88, e92–e95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osterloh, A.; Papp, S.; Moderzynski, K.; Kuehl, S.; Richardt, U.; Fleischer, B. Persisting Rickettsia typhi Causes Fatal Central Nervous System Inflammation. Infect. Immun. 2016, 84, 1615–1632. [Google Scholar] [CrossRef] [Green Version]
- Jay, R.; Armstrong, P.A. Clinical characteristics of Rocky Mountain spotted fever in the United States: A literature review. J. Vector Borne Dis. 2020, 57, 114–120. [Google Scholar] [CrossRef]
- Fisher, J.; Card, G.; Soong, L. Neuroinflammation associated with scrub typhus and spotted fever group rickettsioses. PLoS Negl. Trop. Dis. 2020, 14, e0008675. [Google Scholar] [CrossRef] [PubMed]
- Chiranth, S.B.; Ashwini, K.R.; Gowda, V.K.; Sanjay, K.S.; Ahmed, M.; Basavaraja, G.V. Profile of Neurological Manifestations in Children Presenting with Rickettsial Disease. Indian Pediatr. 2022, 59, 222–225. [Google Scholar] [CrossRef]
- Drexler, N.A.; Dahlgren, F.S.; Heitman, K.N.; Massung, R.F.; Paddock, C.D.; Behravesh, C.B. National Surveillance of Spotted Fever Group Rickettsioses in the United States, 2008-2012. Am. J. Trop. Med. Hyg. 2016, 94, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.H.; Parks, F.M.; Betz, T.G.; Taylor, J.P.; Muehlberger, J.W. Histopathology and immunohistologic demonstration of the distribution of Rickettsia typhi in fatal murine typhus. Am. J. Clin. Pathol. 1989, 91, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.H.; Feng, H.M.; Ladner, S.; Billings, A.N.; Zaki, S.R.; Wear, D.J.; Hightower, B. Immunohistochemical diagnosis of typhus rickettsioses using an anti-lipopolysaccharide monoclonal antibody. Mod. Pathol. 1997, 10, 1038–1042. [Google Scholar] [PubMed]
- Walker, D.H.; Popov, V.L.; Feng, H.M. Establishment of a novel endothelial target mouse model of a typhus group rickettsiosis: Evidence for critical roles for gamma interferon and CD8 T lymphocytes. Lab. Investig. 2000, 80, 1361–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, D.H.; Popov, V.L.; Wen, J.; Feng, H.M. Rickettsia conorii infection of C3H/HeN mice. A model of endothelial-target rickettsiosis. Lab. Investig. 1994, 70, 358–368. [Google Scholar]
- Drevets, D.A.; Leenen, P.J.; Greenfield, R.A. Invasion of the central nervous system by intracellular bacteria. Clin. Microbiol. Rev. 2004, 17, 323–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barichello, T.; Generoso, J.S.; Milioli, G.; Elias, S.G.; Teixeira, A.L. Pathophysiology of bacterial infection of the central nervous system and its putative role in the pathogenesis of behavioral changes. Braz. J. Psychiatry 2013, 35, 81–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baganz, M.D.; Dross, P.E.; Reinhardt, J.A. Rocky Mountain spotted fever encephalitis: MR findings. AJNR Am. J. Neuroradiol. 1995, 16, 919–922. [Google Scholar] [PubMed]
- Joshi, S.G.; Kovács, A.D. Rickettsia rickettsii infection causes apoptotic death of cultured cerebellar granule neurons. J. Med. Microbiol. 2007, 56, 138–141. [Google Scholar] [CrossRef] [Green Version]
- Bohacsova, M.; Filipcik, P.; Opattova, A.; Valarikova, J.; Diaz, M.Q.; Skultety, L.; Novak, M.; Sekeyova, Z. Survival of rat cerebrocortical neurons after rickettsial infection. Microbes Infect. 2015, 17, 845–849. [Google Scholar] [CrossRef]
- Huebner, R.J.; Stamps, P.; Armstrong, C. Rickettsialpox; A newly recognized rickettsial disease; isolation of the etiological agent. Public Health Rep. 1946, 61, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Brezina, R.; Rehácer, J.; Ác, P.; Majerská, M. Two strains of rickettsiae of Rocky Mountain spotted fever group recovered from Dermacentor marginatus ticks in Czechoslovakia. Results of preliminary serological identification. Acta Virol. 1969, 13, 142–145. [Google Scholar] [PubMed]
- Danchenko, M.; Benada, O.; Škultéty, Ľ.; Sekeyová, Z. Culture Isolate of Rickettsia felis from a Tick. Int. J. Env. Res. Public Health 2022, 19, 4321. [Google Scholar] [CrossRef] [PubMed]
- Cente, M.; Mandakova, S.; Filipcik, P. Memantine prevents sensitivity to excitotoxic cell death of rat cortical neurons expressing human truncated tau protein. Cell Mol. Neurobiol. 2009, 29, 945–949. [Google Scholar] [CrossRef]
- Valarikova, J.; Sekeyova, Z.; Skultety, L.; Bohacsova, M.; Quevedo-Diaz, M. New way of purification of pathogenic rickettsiae reducing health risks. Acta Virol. 2016, 60, 206–210. [Google Scholar] [CrossRef] [Green Version]
- Ammerman, N.C.; Beier-Sexton, M.; Azad, A.F. Laboratory maintenance of Rickettsia rickettsii. Curr. Protoc. Microbiol. 2008, 11, 3A.5.1–3A.5.21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wike, D.A.; Tallent, G.; Peacock, M.G.; Ormsbee, R.A. Studies of the rickettsial plaque assay technique. Infect. Immun. 1972, 5, 715–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danchenko, M.; Csaderova, L.; Fournier, P.E.; Sekeyova, Z. Optimized fixation of actin filaments for improved indirect immunofluorescence staining of rickettsiae. BMC Res. Notes 2019, 12, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Brown, K.R.; Otasek, D.; Ali, M.; McGuffin, M.J.; Xie, W.; Devani, B.; Toch, I.L.; Jurisica, I. NAViGaTOR: Network Analysis, Visualization and Graphing Toronto. Bioinformatics 2009, 25, 3327–3329. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, M.J.; Byrge, K.C.; Ivey, K.S.; Pruthi, S.; Bloch, K.C. Meningoencephalitis due to Spotted Fever Rickettsioses, Including Rocky Mountain Spotted Fever. Clin. Infect. Dis. 2020, 71, 188–195. [Google Scholar] [CrossRef]
- Dahlgren, F.S.; Holman, R.C.; Paddock, C.D.; Callinan, L.S.; McQuiston, J.H. Fatal Rocky Mountain spotted fever in the United States, 1999–2007. Am. J. Trop. Med. Hyg. 2012, 86, 713–719. [Google Scholar] [CrossRef] [Green Version]
- Eldin, C.; Parola, P. Rickettsioses as causes of CNS infection in southeast Asia. Lancet Glob. Health 2015, 3, e67–e68. [Google Scholar] [CrossRef] [Green Version]
- Poppell, M.; Hammel, G.; Ren, Y. Immune Regulatory Functions of Macrophages and Microglia in Central Nervous System Diseases. Int. J. Mol. Sci. 2023, 24, 5925. [Google Scholar] [CrossRef]
- Sahni, S.K.; Rydkina, E. Host-cell interactions with pathogenic Rickettsia species. Future Microbiol. 2009, 4, 323–339. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.H.; Ismail, N. Emerging and re-emerging rickettsioses: Endothelial cell infection and early disease events. Nat. Rev. Microbiol. 2008, 6, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Clifton, D.R.; Goss, R.A.; Sahni, S.K.; van Antwerp, D.; Baggs, R.B.; Marder, V.J.; Silverman, D.J.; Sporn, L.A. NF-kappa B-dependent inhibition of apoptosis is essential for host cellsurvival during Rickettsia rickettsii infection. Proc. Natl. Acad. Sci. USA 1998, 95, 4646–4651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.R.; Burkhardt, N.Y.; Kurtti, T.J.; Oliver, J.D.; Price, L.D.; Cull, B.; Thorpe, C.J.; Thiel, M.S.; Munderloh, U.G. Mitochondrion-Dependent Apoptosis Is Essential for Rickettsia parkeri Infection and Replication in Vector Cells. mSystems 2021, 6, 2. [Google Scholar] [CrossRef]
- Mansueto, P.; Vitale, G.; Cascio, A.; Seidita, A.; Pepe, I.; Carroccio, A.; di Rosa, S.; Rini, G.B.; Cillari, E.; Walker, D.H. New insight into immunity and immunopathology of Rickettsial diseases. Clin. Dev. Immunol. 2012, 2012, 967852. [Google Scholar] [CrossRef] [PubMed]
- Jordan, J.M.; Woods, M.E.; Olano, J.; Walker, D.H. The absence of Toll-like receptor 4 signaling in C3H/HeJ mice predisposes them to overwhelming rickettsial infection and decreased protective Th1 responses. Infect. Immun. 2008, 76, 3717–3724. [Google Scholar] [CrossRef] [Green Version]
- Bechelli, J.; Smalley, C.; Zhao, X.; Judy, B.; Valdes, P.; Walker, D.H.; Fang, R. MyD88 Mediates Instructive Signaling in Dendritic Cells and Protective Inflammatory Response during Rickettsial Infection. Infect. Immun. 2016, 84, 883–893. [Google Scholar] [CrossRef] [Green Version]
- Quevedo-Diaz, M.A.; Song, C.; Xiong, Y.; Chen, H.; Wahl, L.M.; Radulovic, S.; Medvedev, A.E. Involvement of TLR2 and TLR4 in cell responses to Rickettsia akari. J. Leukoc. Biol. 2010, 88, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Sahni, A.; Fang, R.; Sahni, S.K.; Walker, D.H. Pathogenesis of Rickettsial Diseases: Pathogenic and Immune Mechanisms of an Endotheliotropic Infection. Annu. Rev. Pathol. 2019, 14, 127–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engstrom, P.; Burke, T.P.; Mitchell, G.; Ingabire, N.; Mark, K.G.; Golovkine, G.; Iavarone, A.T.; Rape, M.; Cox, J.S.; Welch, M.D. Evasion of autophagy mediated by Rickettsia surface protein OmpB is critical for virulence. Nat. Microbiol. 2019, 4, 2538–2551. [Google Scholar] [CrossRef]
- Wang, X.R.; Cull, B. Apoptosis and Autophagy: Current Understanding in Tick-Pathogen Interactions. Front. Cell Infect. Microbiol. 2022, 12, 784430. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cente, M.; Danchenko, M.; Skultety, L.; Filipcik, P.; Sekeyova, Z. Rickettsia Deregulates Genes Coding for the Neurotoxic Cell Response Pathways in Cerebrocortical Neurons In Vitro. Cells 2023, 12, 1235. https://doi.org/10.3390/cells12091235
Cente M, Danchenko M, Skultety L, Filipcik P, Sekeyova Z. Rickettsia Deregulates Genes Coding for the Neurotoxic Cell Response Pathways in Cerebrocortical Neurons In Vitro. Cells. 2023; 12(9):1235. https://doi.org/10.3390/cells12091235
Chicago/Turabian StyleCente, Martin, Monika Danchenko, Ludovit Skultety, Peter Filipcik, and Zuzana Sekeyova. 2023. "Rickettsia Deregulates Genes Coding for the Neurotoxic Cell Response Pathways in Cerebrocortical Neurons In Vitro" Cells 12, no. 9: 1235. https://doi.org/10.3390/cells12091235