
The Mycobacterium tuberculosis PE_PGRS Protein Family Acts as an Immunological Decoy to Subvert Host Immune Response

 ,
, 
Abstract
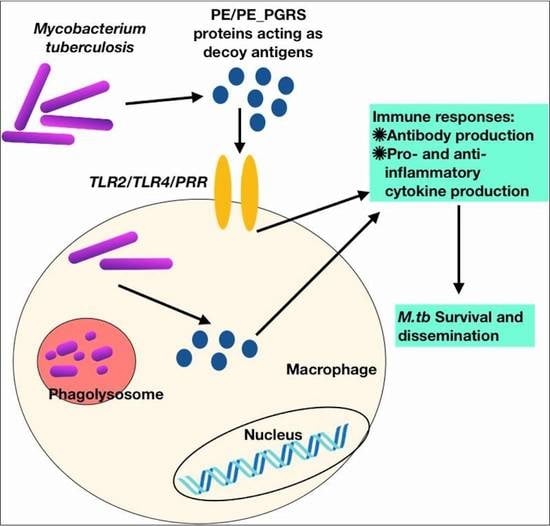
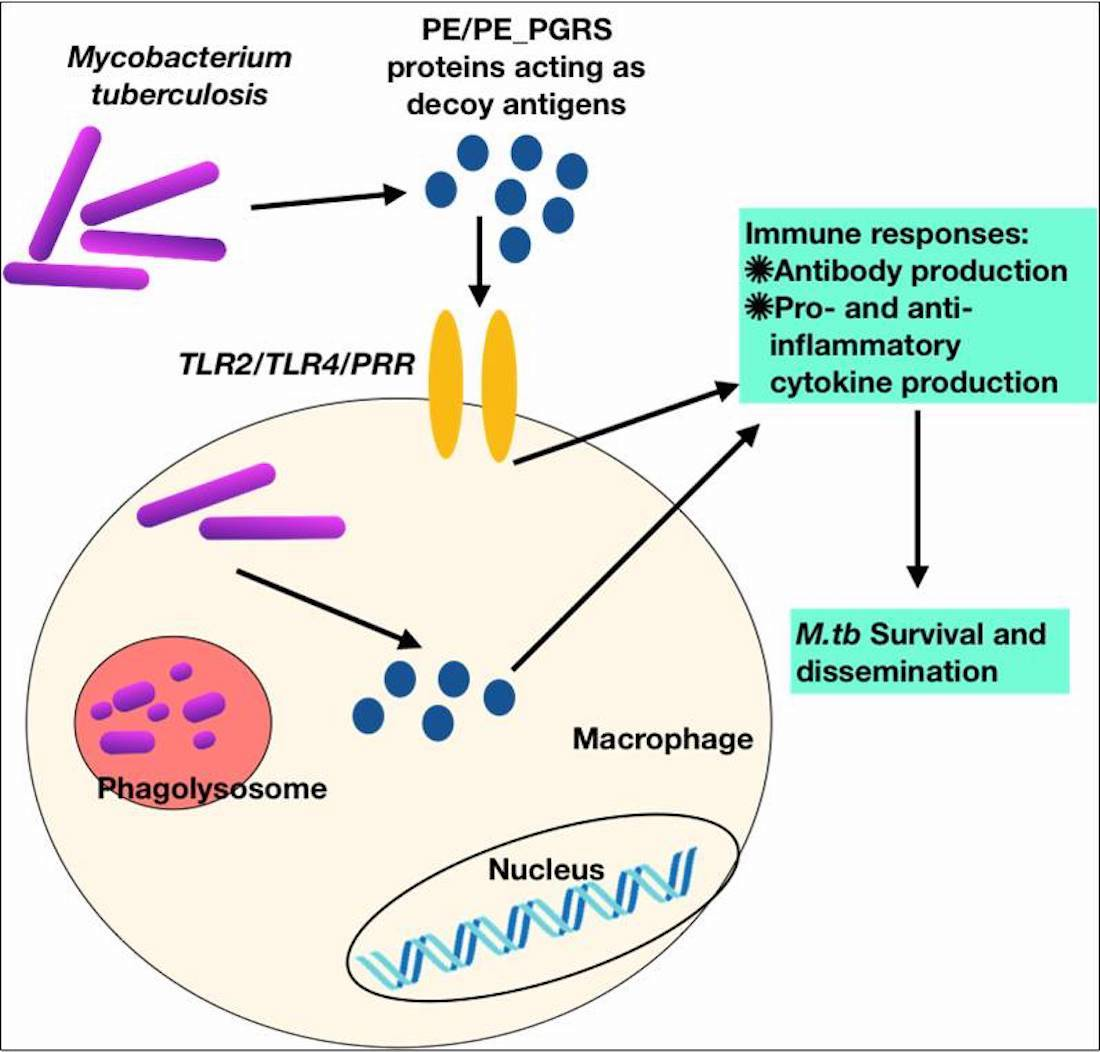
:
1. Introduction
2. Modulators of the Host-Immune Response: PE/PPE Proteins
3. Ambiguous Immune Responses of PE/PPE/PE_PGRS Proteins of M.tb
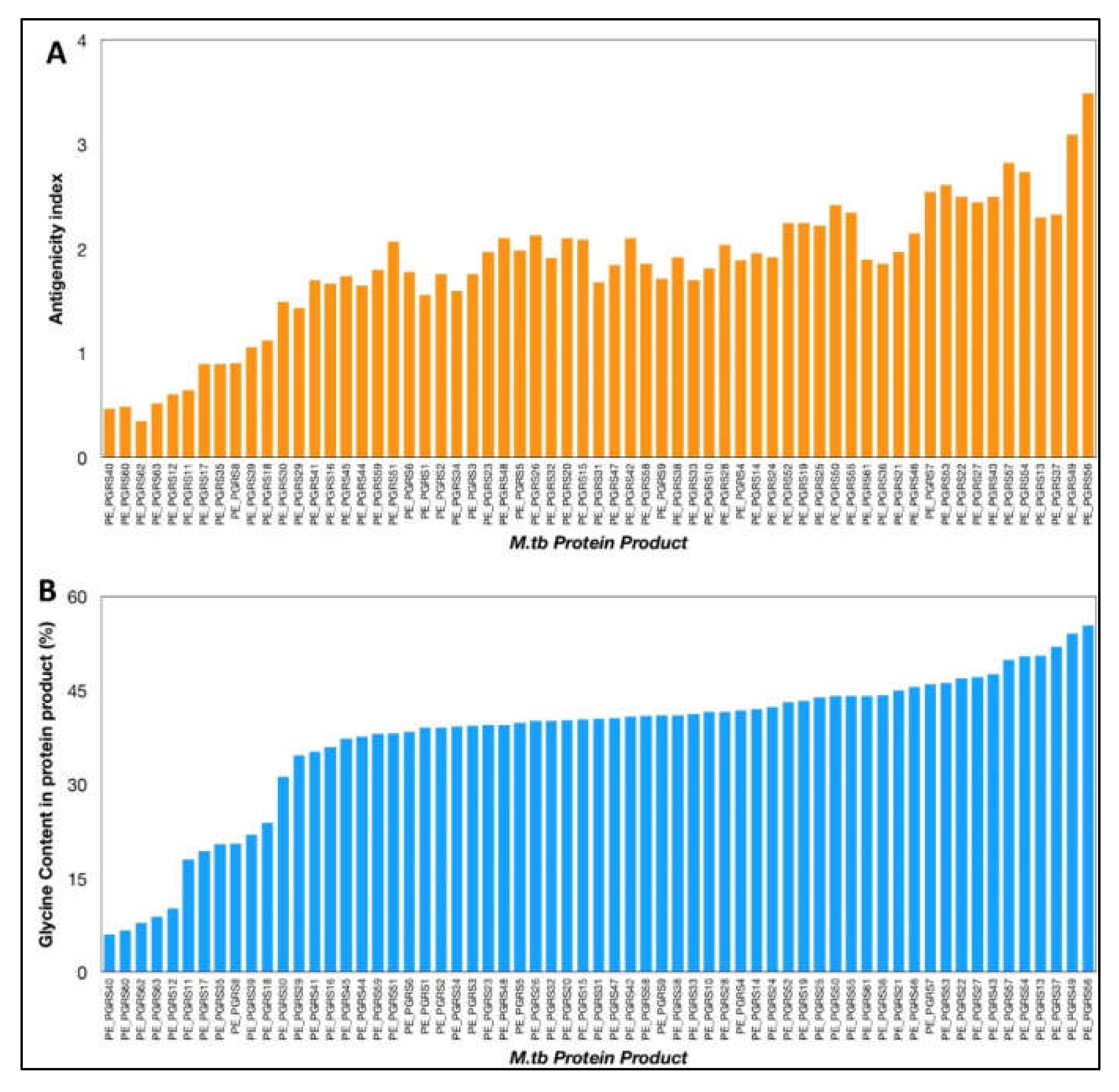
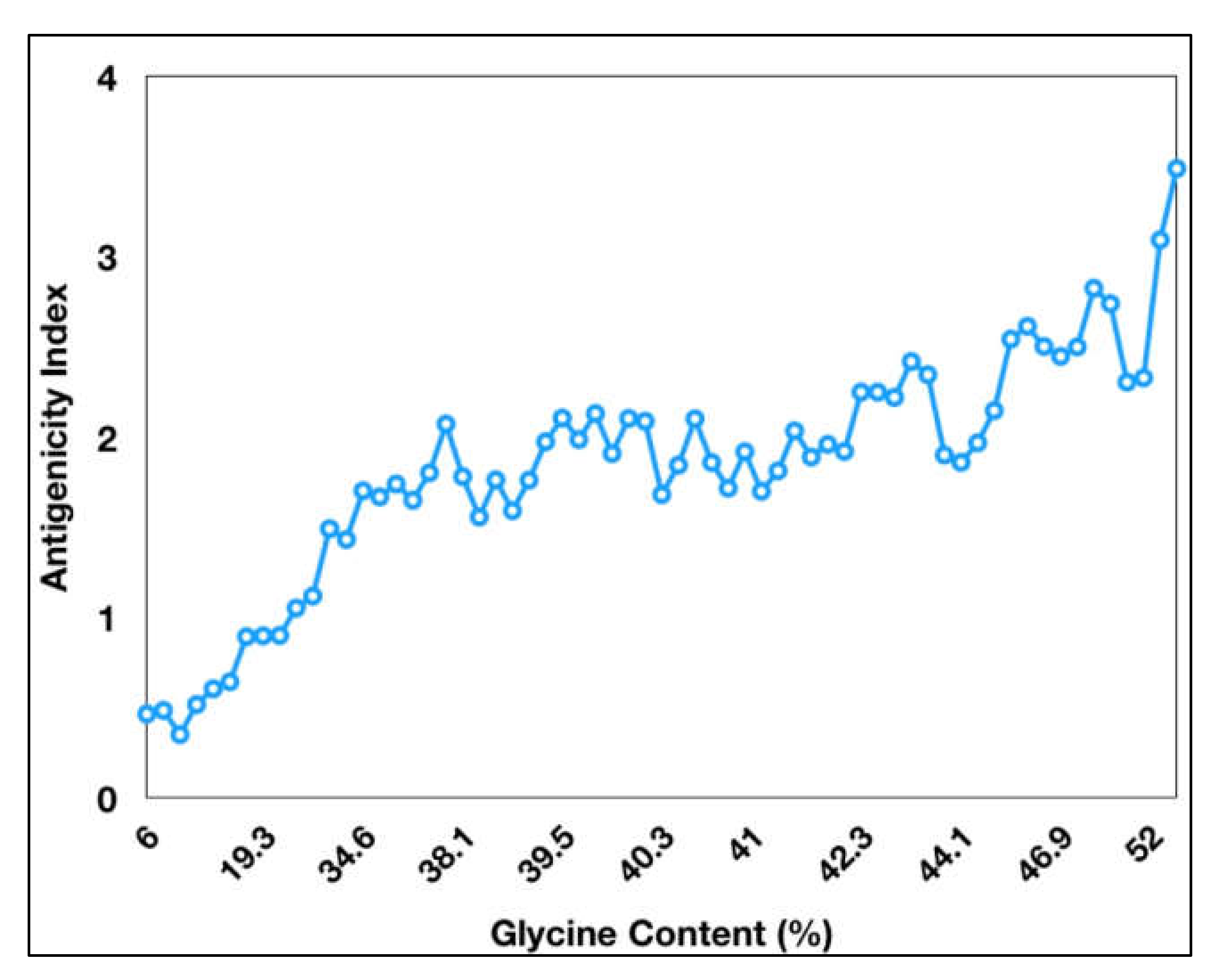
4. Immune Evasion and Subversion Properties of PE_PGRS Proteins: A Possible Reflection of Antigenic Variation, Disordered Nature and Glycine Content
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chakaya, J.; Khan, M.; Ntoumi, F.; Aklillu, E.; Fatima, R.; Mwaba, P.; Kapata, N.; Mfinanga, S.; Hasnain, S.E.; Katoto, P.D.M.C.; et al. Global Tuberculosis Report 2020; Reflections on the Global TB burden, treatment and prevention efforts. Int. J. Infect. Dis. 2021, 11, S1201–9712. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Ehtesham, N.Z.; Hasnain, S.E. Two parallel pandemics: The challenges faced by countries with COVID-19 and TB. Int. J. Tuberc. Lung Dis. 2020, 24, 1319–1320. [Google Scholar] [CrossRef]
- WHO. Global Tuberculosis Report 2019 Geneva. Global Tuberculosis Report 2019; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Ehrt, S.; Schnappinger, D. Mycobacterial survival strategies in the phagosome: Defence against host stresses. Cell. Microbiol. 2009, 11, 1170–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meena, L.S. Interrelation of Ca2+ and PE_PGRS proteins during Mycobacterium tuberculosispathogenesis. J. Biosci. 2019, 44, 24. [Google Scholar] [CrossRef] [PubMed]
- Nair, S. Immunomodulatory Role of Mycobacterial PE/PPE Family of Proteins. Proc. Ind. Natl. Sci. Acad. 2014, 80, 1055. [Google Scholar] [CrossRef]
- Pandey, S.; Tripathi, D.; Khubaib, M.; Kumar, A.; Sheikh, J.A.; Sumanlatha, G.; Ehtesham, N.Z.; Hasnain, S.E. Mycobacterium tuberculosis Peptidyl-Prolyl Isomerases Are Immunogenic, Alter Cytokine Profile and Aid in Intracellular Survival. Front. Cell. Infect. Microbiol. 2017, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Gomes, M.S.; Paul, S.; Moreira, A.L.; Appelberg, R.; Rabinovitch, M.; Kaplan, G. Survival of Mycobacterium avium and Mycobacterium tuberculosis in Acidified Vacuoles of Murine Macrophages. Infect. Immun. 1999, 67, 3199–3206. [Google Scholar] [CrossRef] [Green Version]
- Meena, L.S. Rajni Survival mechanisms of pathogenic Mycobacterium tuberculosis H37Rv. FEBS J. 2010, 277, 2416–2427. [Google Scholar] [CrossRef]
- Baena, A.; Porcelli, S.A. Evasion and subversion of antigen presentation by Mycobacterium tuberculosis. Tissue Antigens 2009, 74, 189–204. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, M.F.; Saini, N.K.; Porcelli, S.A. Evasion of Innate and Adaptive Immunity by Mycobacterium tuberculosis. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Paulus, J.K.; Van Der Hoorn, R.A.L. Tricked or trapped—Two decoy mechanisms in host–pathogen interactions. PLoS Pathog. 2018, 14, e1006761. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Dobrindt, U.; Hacker, J.; Hasnain, S. Genomic fluidity and pathogenic bacteria: Applications in diagnostics, epidemiology and intervention. Nat. Rev. Genet. 2008, 6, 387–394. [Google Scholar] [CrossRef]
- Sampson, S.L. Mycobacterial PE/PPE Proteins at the Host-Pathogen Interface. J. Immunol. Res. 2011, 2011, 497203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Pittius, N.C.G.; Sampson, S.L.; Lee, H.; Kim, Y.; Van Helden, P.D.; Warren, R.M. Evolution and expansion of the Mycobacterium tuberculosis PE and PPE multigene families and their association with the duplication of the ESAT-6 (esx) gene cluster regions. BMC Evol. Biol. 2006, 6, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, J.; Khubaib, M.; Sheikh, J.A.; Pancsa, R.; Kumar, S.; Srinivasan, A.; Babu, M.M.; Hasnain, S.E.; Ehtesham, N.Z. Disorder-to-order transition in PE–PPE proteins of Mycobacterium tuberculosis augments the pro-pathogen immune response. FEBS Open Bio 2019, 10, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Akhter, Y.; Ehebauer, M.T.; Mukhopadhyay, S.; Hasnain, S. The PE/PPE multigene family codes for virulence factors and is a possible source of mycobacterial antigenic variation: Perhaps more? Biochimie 2012, 94, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Delogu, G.; Pusceddu, C.; Bua, A.; Fadda, G.; Brennan, M.J.; Zanetti, S. Rv1818c-encoded PE_PGRS protein of Mycobacterium tuberculosis is surface exposed and influences bacterial cell structure. Mol. Microbiol. 2004, 52, 725–733. [Google Scholar] [CrossRef]
- Cadieux, N.; Parra, M.; Cohen, H.; Maric, D.; Morris, S.L.; Brennan, M.J. Induction of cell death after localization to the host cell mitochondria by the Mycobacterium tuberculosis PE_PGRS33 protein. Microbiology 2011, 157, 793–804. [Google Scholar] [CrossRef] [Green Version]
- Bottai, D.; Di Luca, M.; Majlessi, L.; Frigui, W.; Simeone, R.; Sayes, F.; Bitter, W.; Brennan, M.J.; Leclerc, C.; Batoni, G.; et al. Disruption of the ESX-5 system of Mycobacterium tuberculosis causes loss of PPE protein secretion, reduction of cell wall integrity and strong attenuation. Mol. Microbiol. 2012, 83, 1195–1209. [Google Scholar] [CrossRef] [Green Version]
- Majlessi, L.; Prados-Rosales, R.; Casadevall, A.; Brosch, R. Release of mycobacterial antigens. Immunol. Rev. 2015, 264, 25–45. [Google Scholar] [CrossRef] [PubMed]
- Mohareer, K.; Tundup, S.; Hasnain, S. Transcriptional Regulation of Mycobacterium tuberculosis PE/PPE Genes: A Molecular Switch to Virulence. J. Mol. Microbiol. Biotechnol. 2011, 21, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Balaji, K.N.; Goyal, G.; Narayana, Y.; Srinivas, M.; Chaturvedi, R.; Mohammad, S. Apoptosis triggered by Rv1818c, a PE family gene from Mycobacterium tuberculosis is regulated by mitochondrial intermediates in T cells. Microbes Infect. 2007, 9, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, S.H.E. How can immunology contribute to the control of tuberculosis? Nat. Rev. Immunol. 2001, 1, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Thi, E.P.; Lambertz, U.; Reiner, N.E. Sleeping with the Enemy: How Intracellular Pathogens Cope with a Macrophage Lifestyle. PLoS Pathog. 2012, 8, e1002551. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Pathak, S.K.; Banerjee, A.; Pathak, S.; Bhattacharyya, A.; Yang, Z.; Talarico, S.; Kundu, M.; Basu, J. Execution of Macrophage Apoptosis by PE_PGRS33 of Mycobacterium tuberculosis Is Mediated by Toll-like Receptor 2-dependent Release of Tumor Necrosis Factor-α. J. Biol. Chem. 2007, 282, 1039–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaulieu, A.M.; Rath, P.; Imhof, M.; Siddall, M.E.; Roberts, J.; Schnappinger, D.; Nathan, C.F. Genome-Wide Screen for Mycobacterium tuberculosis Genes That Regulate Host Immunity. PLoS ONE 2010, 5, e15120. [Google Scholar] [CrossRef] [PubMed]
- Bansal, K.; Sinha, A.Y.; Ghorpade, D.S.; Togarsimalemath, S.K.; Patil, S.A.; Kaveri, S.V.; Balaji, K.N.; Bayry, J. Src homology 3-interacting domain of Rv1917c of Mycobacterium tuberculosis induces selective maturation of human dendritic cells by regulating PI3K-MAPK-NF-κB signaling and drives Th2 immune responses. J. Biol. Chem. 2010, 285, 36511–36522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daim, S.; Kawamura, I.; Tsuchiya, K.; Hara, H.; Kurenuma, T.; Shen, Y.; Dewamitta, S.R.; Sakai, S.; Nomura, T.; Qu, H.; et al. Expression of the Mycobacterium tuberculosis PPE37 protein in Mycobacterium smegmatis induces low tumour necrosis factor alpha and interleukin 6 production in murine macrophages. J. Med. Microbiol. 2011, 60, 582–591. [Google Scholar] [CrossRef]
- Tiwari, B.M.; Kannan, N.; Vemu, L.; Raghunand, T.R. The Mycobacterium tuberculosis PE Proteins Rv0285 and Rv1386 Modulate Innate Immunity and Mediate Bacillary Survival in Macrophages. PLoS ONE 2012, 7, e51686. [Google Scholar] [CrossRef] [Green Version]
- Bhat, K.H.; Das, A.; Srikantam, A.; Mukhopadhyay, S. PPE 2 protein of Mycobacterium tuberculosis may inhibit nitric oxide in activated macrophages. Ann. N. Y. Acad. Sci. 2013, 1283, 97–101. [Google Scholar] [CrossRef]
- Singh, V.; Jamwal, S.; Jain, R.; Verma, P.; Gokhale, R.; Rao, K.V. Mycobacterium tuberculosis-Driven Targeted Recalibration of Macrophage Lipid Homeostasis Promotes the Foamy Phenotype. Cell Host Microbe 2012, 12, 669–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, R.K.; Mukhopadhyay, S.; Chakhaiyar, P.; Sharma, N.; Murthy, K.J.R.; Katoch, V.M.; Hasnain, S.E. PPE Antigen Rv2430c of Mycobacterium tuberculosis Induces a Strong B-Cell Response. Infect. Immun. 2003, 71, 6338–6343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tundup, S.; Pathak, N.; Ramanadham, M.; Mukhopadhyay, S.; Murthy, K.J.R.; Ehtesham, N.Z.; Hasnain, S. The Co-Operonic PE25/PPE41 Protein Complex of Mycobacterium tuberculosis Elicits Increased Humoral and Cell Mediated Immune Response. PLoS ONE 2008, 3, e3586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khubaib, M.; Sheikh, J.A.; Pandey, S.; Srikanth, B.; Bhuwan, M.; Khan, N.; Hasnain, S.E.; Ehtesham, N.Z. Mycobacterium tuberculosis Co-operonic PE32/PPE65 Proteins Alter Host Immune Responses by Hampering Th1 Response. Front. Microbiol. 2016, 7, 719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehtram, A.; Shariq, M.; Ali, S.; Ehtesham, N.Z.; Hasnain, S.E. ESX5 associated PE proteins of Mycobacterium tuberculosis drives Th1 type polarization: An immunomodulatory paradigm. Am. Assoc. Immnol. 2020, 12, 2680. [Google Scholar] [CrossRef]
- Li, H.; Li, Q.; Yu, Z.; Zhou, M.; Xie, J. Mycobacterium tuberculosis PE13 (Rv1195) manipulates the host cell fate via p38-ERK-NF-κB axis and apoptosis. Apoptosis 2016, 21, 795–808. [Google Scholar] [CrossRef]
- Deng, W.; Yang, W.; Zeng, J.; Abdalla, A.E.; Xie, J. Mycobacterium tuberculosis PPE32 promotes cytokines production and host cell apoptosis through caspase cascade accompanying with enhanced ER stress response. Oncotarget 2016, 7, 67347–67359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNamara, M.; Tzeng, S.-C.; Maier, C.; Zhang, L.; Bermudez, L.E. Surface proteome of “Mycobacterium avium subsp. hominissuis” during the early stages of macrophage infection. Infect. Immun. 2012, 80, 1868–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Rao, R.N.; Reddy, J.R.C.; Prasad, R.B.N.; Kotturu, S.K.; Ghosh, S.; Mukhopadhyay, S. PE11, a PE/PPE family protein of Mycobacterium tuberculosis is involved in cell wall remodeling and virulence. Sci. Rep. 2016, 6, 21624. [Google Scholar] [CrossRef] [Green Version]
- Iantomasi, R.; Sali, M.; Cascioferro, A.; Palucci, I.; Zumbo, A.; Soldini, S.; Rocca, S.; Greco, E.; Maulucci, G.; De Spirito, M.; et al. PE_PGRS30 is required for the full virulence of Mycobacterium tuberculosis. Cell. Microbiol. 2011, 14, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Wang, D.; Li, M.; Wang, H.; Yu, J.; Wang, C.; Liu, J.; Gao, Q. PPE38 Modulates the Innate Immune Response and Is Required for Mycobacterium marinum Virulence. Infect. Immun. 2011, 80, 43–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilo, J.; Alonso, H.; Uranga, S.; Marinova, D.; Arbues, A.; De Martino, A.; Anel, A.; Monzon, M.; Badiola, J.; Pardo, J. ESX-1-induced apoptosis is involved in cell-to-cell spread of M ycobacterium tuberculosis. Cell. Microbiol. 2013, 15, 1994–2005. [Google Scholar] [CrossRef]
- Grover, S.; Sharma, T.; Singh, Y.; Kohli, S.; Manjunath, P.; Singh, A.; Semmler, T.; Wieler, L.H.; Tedin, K.; Ehtesham, N.Z.; et al. The PGRS Domain of Mycobacterium tuberculosis PE_PGRS Protein Rv0297 Is Involved in Endoplasmic Reticulum Stress-Mediated Apoptosis through Toll-Like Receptor 4. mBio 2018, 9, e01017-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, J.; Farhana, A.; Pancsa, R.; Arora, S.K.; Srinivasan, A.; Tyagi, A.K.; Babu, M.M.; Ehtesham, N.Z.; Hasnain, S.E. Contrasting Function of Structured N-Terminal and Unstructured C-Terminal Segments of Mycobacterium tuberculosis PPE37 Protein. mBio 2018, 9, e01712-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tundup, S.; Mohareer, K.; Hasnain, S.E. Mycobacterium tuberculosisPE25/PPE41 protein complex induces necrosis in macrophages: Role in virulence and disease reactivation? FEBS Open Bio 2014, 4, 822–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dheenadhayalan, V.; Delogu, G.; Brennan, M.J. Expression of the PE_PGRS 33 protein in Mycobacterium smegmatis triggers necrosis in macrophages and enhanced mycobacterial survival. Microbes Infect. 2006, 8, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Kohli, S.; Singh, Y.; Sharma, K.; Mittal, A.; Ehtesham, N.Z.; Hasnain, S.E. Comparative genomic and proteomic analyses of PE/PPE multigene family of Mycobacterium tuberculosis H37Rv and H37Ra reveal novel and interesting differences with implications in virulence. Nucl. Acids Res. 2012, 40, 7113–7122. [Google Scholar] [CrossRef] [PubMed]
- Bansal, K.; Elluru, S.R.; Narayana, Y.; Chaturvedi, R.; Patil, S.A.; Kaveri, S.V.; Bayry, J.; Balaji, K.N. PE_PGRS Antigens of Mycobacterium tuberculosisInduce Maturation and Activation of Human Dendritic Cells. J. Immunol. 2010, 184, 3495–3504. [Google Scholar] [CrossRef] [Green Version]
- Sharma, T.; Grover, S.; Arora, N.; Manjunath, P.; Ehtesham, N.Z.; Hasnain, S.E. PGRS Domain of Rv0297 of Mycobacterium tuberculosis Is Involved in Modulation of Macrophage Functions to Favor Bacterial Persistence. Front. Cell. Infect. Microbiol. 2020, 10, 451. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.; Singh, J.; Grover, S.; Manjunath, P.; Firdos, F.; Alam, A.; Ehtesham, N.Z.; Hasnain, S.E. PGRS Domain of Rv0297 of Mycobacterium tuberculosis Functions in A Calcium Dependent Manner. Int. J. Mol. Sci. 2021, 22, 9390. [Google Scholar] [CrossRef]
- Palucci, I.; Camassa, S.; Cascioferro, A.; Sali, M.; Anoosheh, S.; Zumbo, A.; Minerva, M.; Iantomasi, R.; De Maio, F.; Di Sante, G.; et al. PE_PGRS33 Contributes to Mycobacterium tuberculosis Entry in Macrophages through Interaction with TLR2. PLoS ONE 2016, 11, e0150800. [Google Scholar] [CrossRef] [PubMed]
- Delogu, G.; Brennan, M.J. Comparative Immune Response to PE and PE_PGRS Antigens of Mycobacterium tuberculosis. Infect. Immun. 2001, 69, 5606–5611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, I.; Parada, C.; Acosta-Gão, E.; Espitia, C. The PGRS Domain from PE_PGRS33 of Mycobacterium tuberculosis is Target of Humoral Immune Response in Mice and Humans. Front. Immunol. 2014, 5, 236. [Google Scholar] [CrossRef]
- Singh, P.P.; Parra, M.; Cadieux, N.; Brennan, M.J. A comparative study of host response to three Mycobacterium tuberculosis PE_PGRS proteins. Microbiology 2008, 154, 3469–3479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Li, W.; Zeng, J.; Zhao, Q.; Li, C.; Zhao, Y.; Xie, J. Mycobacterium tuberculosis PPE family protein Rv1808 manipulates cytokines profile via co-activation of MAPK and NF-κB signaling pathways. Cell. Physiol. Biochem. 2014, 33, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Medha, L.S.; Sharma, S.; Sharma, M. Proline-Glutamate/Proline-Proline-Glutamate (PE/PPE) proteins of Mycobacterium tuberculosis: The multifaceted immune-modulators. Acta Trop. 2021, 222, 106035. [Google Scholar] [CrossRef]
- Sharma, N.; Shariq, M.; Quadir, N.; Singh, J.; Sheikh, J.A.; Hasnain, S.E.; Ehtesham, N.Z. Mycobacterium tuberculosis Protein PE6 (Rv0335c), a Novel TLR4 Agonist, Evokes an Inflammatory Response and Modulates the Cell Death Pathways in Macrophages to Enhance Intracellular Survival. Front. Immunol. 2021, 12, 696491. [Google Scholar] [CrossRef] [PubMed]
- Meena, L.S. An overview to understand the role of PE_PGRS family proteins inMycobacterium tuberculosisH37Rv and their potential as new drug targets. Biotechnol. Appl. Biochem. 2014, 62, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Tripathi, D.K.; Singh, P.K.; Sharma, S.; Srivastava, K.K. Protective and survival efficacies of Rv0160c protein in murine model of Mycobacterium tuberculosis. Appl. Microbiol. Biotechnol. 2012, 97, 5825–5837. [Google Scholar] [CrossRef] [PubMed]
- Chatrath, S.; Gupta, V.K.; Dixit, A.; Garg, L.C. The Rv1651c-encoded PE-PGRS30 protein expressed in Mycobacterium smegmatis exhibits polar localization and modulates its growth profile. FEMS Microbiol. Lett. 2011, 322, 194–199. [Google Scholar] [CrossRef] [Green Version]
- Chatrath, S.; Gupta, V.K.; Dixit, A.; Garg, L.C. PE_PGRS30 of Mycobacterium tuberculosis mediates suppression of proinflammatory immune response in macrophages through its PGRS and PE domains. Microbes Infect. 2016, 18, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Gallant, J.; Heunis, T.; Beltran, C.; Schildermans, K.; Bruijns, S.; Mertens, I.; Bitter, W.; Sampson, S.L. PPE38-Secretion-Dependent Proteins of M. tuberculosis Alter NF-kB Signalling and Inflammatory Responses in Macrophages. Front. Immunol. 2021, 12, 2440. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Ramaswamy, P.A.; Ghosh, S.; Joshi, D.; Pathak, N.; Siddiqui, I.; Sharma, P.; Hasnain, S.; Mande, S.C.; Mukhopadhyay, S. The PPE18 ofMycobacterium tuberculosisInteracts with TLR2 and Activates IL-10 Induction in Macrophage. J. Immunol. 2009, 183, 6269–6281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, S.; Pandey, A.D.; Mukhopadhyay, S. The PPE18 Protein ofMycobacterium tuberculosisInhibits NF-κB/rel–Mediated Proinflammatory Cytokine Production by Upregulating and Phosphorylating Suppressor of Cytokine Signaling 3 Protein. J. Immunol. 2011, 186, 5413–5424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakhaiyar, P.; Nagalakshmi, Y.; Aruna, B.; Murthy, K.J.R.; Katoch, V.M.; Hasnain, S.E. Regions of High Antigenicity within the Hypothetical PPE Major Polymorphic Tandem Repeat Open-Reading Frame, Rv2608, Show a Differential Humoral Response and a Low T Cell Response in Various Categories of Patients with Tuberculosis. J. Infect. Dis. 2004, 190, 1237–1244. [Google Scholar] [CrossRef]
- Huang, Y.; Zhou, X.; Bai, Y.; Yang, L.; Yin, X.; Wang, Z.; Zhao, D. Phagolysosome maturation of macrophages was reduced by PE_PGRS 62 protein expressing in Mycobacterium smegmatis and induced in IFN-γ priming. Vet. Microbiol. 2012, 160, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Thi, E.P.; Hong, C.J.H.; Sanghera, G.; Reiner, N.E. Identification of theMycobacterium tuberculosisprotein PE-PGRS62 as a novel effector that functions to block phagosome maturation and inhibit iNOS expression. Cell. Microbiol. 2012, 15, 795–808. [Google Scholar] [CrossRef]
- Koh, K.W.; Soh, S.E.; Seah, G.T. Strong Antibody Responses to Mycobacterium tuberculosis PE-PGRS62 Protein Are Associated with Latent and Active Tuberculosis. Infect. Immun. 2009, 77, 3337–3343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, B.; Sorry, A.; Raghunand, T.R. An immunomodulatory role for the M ycobacterium tuberculosis region of difference 1 locus proteins PE 35 (R v3872) and PPE 68 (R v3873). FEBS J. 2014, 281, 1556–1570. [Google Scholar] [CrossRef]
- Tundup, S.; Akhter, Y.; Thiagarajan, D.; Hasnain, S. Clusters of PE and PPE genes ofMycobacterium tuberculosisare organized in operons: Evidence that PE Rv2431c is co-transcribed with PPE Rv2430c and their gene products interact with each other. FEBS Lett. 2006, 580, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Riley, R.; Pellegrini, M.; Eisenberg, D. Identifying cognate binding pairs among a large set of paralogs: The case of PE/PPE Proteins of Mycobacterium tuberculosis. PLoS Commpuattional Biol. 2008, 4, e1000174. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, B.; Ramakrishnan, U.M.; Raghunand, T.R. The Mycobacterium tuberculosis protein pair PE9 (Rv1088)-PE10 (Rv1089) forms heterodimers and induces macrophage apoptosis through Toll-like receptor 4. Cell. Microbiol. 2015, 17, 1653–1669. [Google Scholar] [CrossRef] [PubMed]
- Ehtram, A.; Shariq, M.; Ali, S.; Quadir, N.; Sheikh, J.A.; Ahmad, F.; Sharma, T.; Ehtesham, N.Z.; Hasnain, S.E. Teleological cooption of Mycobacterium tuberculosis PE/PPE proteins as porins: Role in molecular immigration and emigration. Int. J. Med. Microbiol. 2021, 311, 151495. [Google Scholar] [CrossRef] [PubMed]
- Delogu, G.; Bigi, F.; Hasnain, S.E.; Cataldi, A. Enigmatic Proteins from the Surface: The Erp, PE, and PPE Protein Families; Wiley: Hoboken, NJ, USA, 2014; pp. 133–151. [Google Scholar] [CrossRef]
- Bowie, A.G.; Unterholzner, L. Viral evasion and subversion of pattern-recognition receptor signalling. Nat. Rev. Immunol. 2008, 8, 911–922. [Google Scholar] [CrossRef]
- Finlay, B.B.; McFadden, G. Anti-Immunology: Evasion of the Host Immune System by Bacterial and Viral Pathogens. Cell 2006, 124, 767–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoniou, A.N.; Powis, S. Pathogen evasion strategies for the major histocompatibility complex class I assembly pathway. Immunology 2008, 124, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Woolard, M.D.; Frelinger, J.A. Outsmarting the host: Bacteria modulating the immune response. Immunol. Res. 2008, 41, 188–202. [Google Scholar] [CrossRef]
- Yu, X.; Feng, J.; Huang, L.; Gao, H.; Liu, J.; Bai, S.; Wu, B.; Xie, J. Molecular Basis Underlying Host Immunity Subversion by Mycobacterium tuberculosis PE/PPE Family Molecules. DNA Cell Biol. 2019, 38, 1178–1187. [Google Scholar] [CrossRef] [PubMed]
- Koh, K.W.; Lehming, N.; Seah, G.T. Degradation-resistant protein domains limit host cell processing and immune detection of mycobacteria. Mol. Immunol. 2009, 46, 1312–1318. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Jr., W.J.S.; Radivojac, P.; Dunker, A.K.; Uversky, V.N. Intrinsic disorder in pathogenic and non-pathogenic microbes: Discovering and analyzing the unfoldomes of early-branching eukaryotes. Mol. BioSyst. 2008, 4, 328–340. [Google Scholar] [CrossRef]
- Tompa, P. Intrinsically unstructured proteins. Trends Biochem. Sci. 2002, 27, 527–533. [Google Scholar] [CrossRef]
- Via, A.; Uyar, B.; Brun, C.; Zanzoni, A. How pathogens use linear motifs to perturb host cell networks. Trends Biochem. Sci. 2015, 40, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Van Der Lee, R.; Buljan, M.; Lang, B.; Weatheritt, R.J.; Daughdrill, G.W.; Dunker, A.K.; Fuxreiter, M.; Gough, J.; Gsponer, J.; Jones, D.T.; et al. Classification of Intrinsically Disordered Regions and Proteins. Chem. Rev. 2013, 114, 6589–6631. [Google Scholar] [CrossRef] [PubMed]
- Davey, N.E.; Travé, G.; Gibson, T.J. How viruses hijack cell regulation. Trends Biochem. Sci. 2011, 36, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Blundell, T.L.; Gupta, M.N.; Hasnain, S.E. Intrinsic disorder in proteins: Relevance to protein assemblies, drug design and host-pathogen interactions. Prog. Biophys. Mol. Biol. 2020, 156, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Ehtesham, N.Z.; Ahmad, J.; Farhan, A.; Khubaib, M.; Kaur, S.; Pancsa, R.; Srinivasan, A.; Kumar, S.; Babu, M.; Hasnain, S.E. Intrinsically Disordered Regions/Proteins Compensate for Genomic Economization In Mycobacterium tuberculosis. FASEB J. 2018, 32, 526.23. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr.; Travers, P.M.W. Pathogens have evolved various means of evading or subverting normal host defenses. In Immunobiology: The Immune System in Health and Disease, Chapter 11, 5th ed.; Garland Science: New York, NY. USA, 2001. [Google Scholar]
- Mohan, G.S.; Li, W.; Ye, L.; Compans, R.W.; Yang, C. Antigenic Subversion: A Novel Mechanism of Host Immune Evasion by Ebola Virus. PLoS Pathog. 2012, 8, e1003065. [Google Scholar] [CrossRef] [Green Version]
- Yeruva, V.C.; Kulkarni, A.; Khandelwal, R.; Sharma, Y.; Raghunand, T.R. The PE_PGRS Proteins of Mycobacterium tuberculosis Are Ca2+ Binding Mediators of Host–Pathogen Interaction. Biochemistry 2016, 55, 4675–4687. [Google Scholar] [CrossRef]
- Ramakrishnan, L.; Federspiel, N.A.; Falkow, S. Granuloma-Specific Expression of Mycobacterium Virulence Proteins from the Glycine-Rich PE-PGRS Family. Science 2000, 288, 1436–1439. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Wheeler, M.D.; Li, X.; Froh, M.; Schemmer, P.; Yin, M.; Bunzendaul, H.; Bradford, B.; Lemasters, J.J. L-Glycine: A novel antiinflammatory, immunomodulatory, and cytoprotective agent. Curr. Opin. Clin. Nutr. Metab. Care 2003, 6, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.T.; Brosch, R.; Parkhill, J.; Garnier, T.; Churcher, C.; Harris, D.; Gordon, S.V.; Eiglmeier, K.; Gas, S.; Barry, C.E.; et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 1998, 396, 190. [Google Scholar] [CrossRef]
- Harding, C.V.; Boom, W.H. Regulation of antigen presentation by Mycobacterium tuberculosis: A role for Toll-like receptors. Nat. Rev. Genet. 2010, 8, 296–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grotzke, J.E.; Siler, A.C.; Lewinsohn, D.A.; Lewinsohn, D.M. Secreted Immunodominant Mycobacterium tuberculosis Antigens Are Processed by the Cytosolic Pathway. J. Immunol. 2010, 185, 4336–4343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bettencourt, P.; Müller, J.; Nicastri, A.; Cantillon, D.; Madhavan, M.; Charles, P.D.; Fotso, C.B.; Wittenberg, R.; Bull, N.; Pinpathomrat, N.; et al. Identification of antigens presented by MHC for vaccines against tuberculosis. Vaccines 2020, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Winslow, G.M.; Cooper, A.; Reiley, W.; Chatterjee, M.; Woodland, D.L. Early T-cell responses in tuberculosis immunity. Immunol. Rev. 2008, 225, 284–299. [Google Scholar] [CrossRef]
- Bold, T.D.; Banaei, N.; Wolf, A.J.; Ernst, J.D. Suboptimal Activation of Antigen-Specific CD4+ Effector Cells Enables Persistence of M. tuberculosis In Vivo. PLoS Pathog. 2011, 7, e1002063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arlehamn, C.S.L.; Gerasimova, A.; Mele, F.; Henderson, R.; Swann, J.; Greenbaum, J.A.; Kim, Y.; Sidney, J.; James, E.A.; Taplitz, R.; et al. Memory T Cells in Latent Mycobacterium tuberculosis Infection Are Directed against Three Antigenic Islands and Largely Contained in a CXCR3+CCR6+ Th1 Subset. PLoS Pathog. 2013, 9, e1003130. [Google Scholar] [CrossRef]
- Dheenadhayalan, V.; Delogu, G.; Sanguinetti, M.; Fadda, G.; Brennan, M.J. Variable Expression Patterns of Mycobacterium tuberculosis PE_PGRS Genes: Evidence that PE_PGRS16 and PE_PGRS26 Are Inversely Regulated In Vivo. J. Bacteriol. 2006, 188, 3721–3725. [Google Scholar] [CrossRef] [Green Version]
- Dillon, D.C.; Alderson, M.R.; Day, C.H.; Lewinsohn, D.; Coler, R.; Bement, T.; Campos-Neto, A.; Skeiky, Y.A.W.; Orme, I.M.; Roberts, A.; et al. Molecular Characterization and Human T-Cell Responses to a Member of a Novel Mycobacterium tuberculosis mtb39 Gene Family. Infect. Immun. 1999, 67, 2941–2950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonanni, D.; Rindi, L.; Lari, N.; Garzelli, C. Immunogenicity of mycobacterial PPE44 (Rv2770c) in Mycobacterium bovis BCG-infected mice. J. Med. Microbiol. 2005, 54, 443–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ates, L.S.; Sayes, F.; Frigui, W.; Ummels, R.; Damen, M.P.M.; Bottai, D.; Behr, M.A.; van Heijst, J.W.J.; Bitter, W.; Majlessi, L.; et al. RD5-mediated lack of PE_PGRS and PPE-MPTR export in BCG vaccine strains results in strong reduction of antigenic repertoire but little impact on protection. PLoS Pathog. 2018, 14, e1007139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abo-Kadoum, M.; Assad, M.; Ali, K.; Uae, M.; Nzaou, S.A.; Gong, Z.; Moaaz, A.; Lambert, N.; Eltoukhy, A.; Xie, J. Mycobacterium tuberculosis PE17 (Rv1646) promotes host cell apoptosis via host chromatin remodeling mediated by reduced H3K9me3 occupancy. Microb. Pathog. 2021, 159, 105147. [Google Scholar] [CrossRef] [PubMed]
- Yi, F.; Hu, J.; Zhu, X.; Wang, Y.; Yu, Q.; Deng, J.; Huang, X.; Ma, Y.; Xie, Y. Transcriptional Profiling of Human Peripheral Blood Mononuclear Cells Stimulated by Mycobacterium tuberculosis PPE57 Identifies Characteristic Genes Associated with Type I Interferon Signaling. Front. Cell. Infect. Microbiol. 2021, 11, 762. [Google Scholar] [CrossRef] [PubMed]
- Ali, K.; Zhen, G.; Nzungize, L.; Stojkoska, A.; Duan, X.; Li, C.; Duan, W.; Xu, J.; Xie, J. Mycobacterium tuberculosis PE31 (Rv3477) Attenuates Host Cell Apoptosis and Promotes Recombinant, M. smegmatis Intracellular Survival via Up-regulating GTPase Guanylate Binding Protein-1. Front. Cell. Infect. Microbiol. 2020, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.S.; Kim, J.-S.; Bin Cha, S.; Kim, S.J.; Kim, H.; Kwon, K.W.; Han, S.J.; Choi, S.Y.; Shin, S.J. Mycobacterium tuberculosis PE27 activates dendritic cells and contributes to Th1-polarized memory immune responses during in vivo infection. Immunobiology 2015, 221, 440–453. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Zeng, J.; Xiang, X.; Li, P.; Xie, J. PE11 (Rv1169c) selectively alters fatty acid components of Mycobacterium smegmatis and host cell interleukin-6 level accompanied with cell death. Front. Microbiol. 2015, 6, 613. [Google Scholar] [CrossRef] [PubMed]
- Dolasia, K.; Nazar, F.; Mukhopadhyay, S. Mycobacterium tuberculosis PPE18 protein inhibits MHC class II antigen presentation and B cell response in mice. Eur. J. Immunol. 2020, 51, 603–619. [Google Scholar] [CrossRef]
- Qureshi, R.; Rameshwaram, N.R.; Battu, M.B.; Mukhopadhyay, S. PPE65 of M. tuberculosis regulate pro-inflammatory signalling through LRR domains of Toll like receptor-2. Biochem. Biophys. Res. Commun. 2018, 508, 152–158. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, E.; Huang, Q.; Ni, W.; Kong, C.; Liu, G.; Li, G.; Su, H.; Wang, H. PPE57 induces activation of macrophages and drives Th1-type immune responses through TLR2. J. Mol. Med. 2015, 93, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Kong, C.; Zhu, L.; Huang, Q.; Luo, L.; Wang, H.; Xu, Y. PPE26 induces TLR2-dependent activation of macrophages and drives Th1-type T-cell immunity by triggering the cross-talk of multiple pathways involved in the host response. Oncotarget 2015, 6, 38517–38537. [Google Scholar] [CrossRef] [Green Version]
- Gong, Z.; Kuang, Z.; Li, H.; Li, C.; Ali, M.K.; Huang, F.; Li, P.; Li, Q.; Huang, X.; Ren, S. Regulation of host cell pyroptosis and cytokines production by Mycobacterium tuberculosis effector PPE60 requires LUBAC mediated NF-κB signaling. Cell. Immunol. 2019, 335, 41–50. [Google Scholar] [CrossRef]
- Su, H.; Zhang, Z.; Liu, Z.; Peng, B.; Kong, C.; Wang, H.; Zhang, Z.; Xu, Y. Mycobacterium tuberculosis PPE60 antigen drives Th1/Th17 responses via Toll-like receptor 2–dependent maturation of dendritic cells. J. Biol. Chem. 2018, 293, 10287–10302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Luo, T.; Zhai, X.; Zhang, C.; Suo, J.; Ma, P.; Wang, C.; Bao, L. PPE11 of Mycobacterium tuberculosis can alter host inflammatory response and trigger cell death. Microb. Pathog. 2018, 126, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Luo, T.; Sun, C.; Yuan, J.; Peng, X.; Zhang, C.; Zhai, X.; Bao, L. PPE27 in Mycobacterium smegmatis Enhances Mycobacterial Survival and Manipulates Cytokine Secretion in Mouse Macrophages. J. Interf. Cytokine Res. 2017, 37, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhang, C.; Zhou, M.; Li, Q.; Li, H.; Duan, W.; Li, X.; Feng, Y.; Xie, J. Mycobacterium tuberculosis PPE44 (Rv2770c) is involved in response to multiple stresses and promotes the macrophage expression of IL-12 p40 and IL-6 via the p38, ERK, and NF-κB signaling axis. Int. Immunopharmacol. 2017, 50, 319–329. [Google Scholar] [CrossRef]
- Asaad, M.; Ali, M.K.; Abo-Kadoum, M.; Lambert, N.; Gong, Z.; Wang, H.; Uae, M.; Nazou, S.A.; Kuang, Z.; Xie, J. Mycobacterium tuberculosis PPE10 (Rv0442c) alters host cell apoptosis and cytokine profile via linear ubiquitin chain assembly complex HOIP-NF-κB signaling axis. Int. Immunopharmacol. 2021, 94, 107363. [Google Scholar] [CrossRef]
- Deng, W.; Long, Q.; Zeng, J.; Li, P.; Yang, W.; Chen, X.; Xie, J. Mycobacterium tuberculosis PE_PGRS41 Enhances the Intracellular Survival of M. smegmatis within Macrophages Via Blocking Innate Immunity and Inhibition of Host Defense. Sci. Rep. 2017, 7, 46716. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Deng, W.; Zeng, J.; Ren, S.; Ali, K.; Gu, Y.; Li, Y.; Xie, J. Mycobacterium tuberculosis PE_PGRS18 enhances the intracellular survival of M. smegmatis via altering host macrophage cytokine profiling and attenuating the cell apoptosis. Apoptosis 2016, 22, 502–509. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | PE Proteins | Role in Immune Modulation | Reference |
|---|---|---|---|
| 1. | PE17 |
| [105,106] |
| |||
| 2. | PE6 |
| [58,105] |
| 3. | PE31 |
| [58,107] |
| 4. | PE13 |
| [37] |
| 5. | PE27 |
| [108] |
| 6. | PE11 |
| [109] |
| 7. | PE5 |
| [30] |
| 8. | PE15 |
| [30] |
| Sr. No. | PPE Proteins | Role in Immune Modulation | Reference |
|---|---|---|---|
| 1. | PPE18 |
| [110] |
| 2. | PPE65 |
| [111] |
| 3. | PPE57 |
| [112] |
| 4. | PPE26 |
| [113] |
| 5. | PPE60 |
| [114,115] |
| 6. | PPE11 |
| [116] |
| 7. | PPE27 |
| [117] |
| 8. | PPE44 |
| [118] |
| 9. | PPE38 |
| [63] |
| 10. | PPE10 |
| [119] |
| 11. | PPE32 |
| [38] |
| 12. | PPE57 |
| [106] |
| Sr. No. | PE_PGRS Proteins | Role in Immune Modulation | Reference |
|---|---|---|---|
| 1. | PE_PGRS41 |
| [120] |
| 2. | PE_PGRS18 |
| [121] |
| 3. | PE_PGRS5 |
| [44,50] |
| 4. | PE_PGRS11 |
| [49] |
| 5. | PE_PGRS17 |
| [49] |
| 6. | PE_PGRS33 |
| [26] |
| 7. | PE_PGRS62 |
| [69] |
| Sr. No. | PE/PPE Proteins | Role in Immune Modulation | Reference |
|---|---|---|---|
| 1. | PE32/PPE65 |
| [35] |
| 2. | PE9/PE10 |
| [72] |
| 3. | PE25/PPE41 |
| [46] |
| 4. | PE35/PPE68 |
| [70] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, T.; Alam, A.; Ehtram, A.; Rani, A.; Grover, S.; Ehtesham, N.Z.; Hasnain, S.E. The Mycobacterium tuberculosis PE_PGRS Protein Family Acts as an Immunological Decoy to Subvert Host Immune Response. Int. J. Mol. Sci. 2022, 23, 525. https://doi.org/10.3390/ijms23010525
Sharma T, Alam A, Ehtram A, Rani A, Grover S, Ehtesham NZ, Hasnain SE. The Mycobacterium tuberculosis PE_PGRS Protein Family Acts as an Immunological Decoy to Subvert Host Immune Response. International Journal of Molecular Sciences. 2022; 23(1):525. https://doi.org/10.3390/ijms23010525
Chicago/Turabian StyleSharma, Tarina, Anwar Alam, Aquib Ehtram, Anshu Rani, Sonam Grover, Nasreen Z. Ehtesham, and Seyed E. Hasnain. 2022. "The Mycobacterium tuberculosis PE_PGRS Protein Family Acts as an Immunological Decoy to Subvert Host Immune Response" International Journal of Molecular Sciences 23, no. 1: 525. https://doi.org/10.3390/ijms23010525